Hannah van Susteren1,*, Guillaume Girier
2,*, Michel J.A.M. van Putten
3,4 , Jaroslav Hlinka
2, Helmut Schmidt
2, Hil G.E. Meijer
1
1 Department of Applied Mathematics, University of Twente, Enschede, the Netherlands
2 Institute of Computer Science, Czech Academy of Sciences, Prague, Czech Republic
3 Department of Neurology and Clinical Neurophysiology, University of Twente, Enschede, the Netherlands
4 Medisch Spectrum Twente, Enschede, the Netherlands
* These authors contributed equally to this work.
Email:
[email protected]
IntroductionEpilepsy is among the most prevalent neurological disorders. The astrocytic excitatory amino acid transporter (EAAT2) plays a key role in regulating excitability, by controlling extracellular glutamate levels and glutamate receptor activation [1,2]. Reduced EAAT2 expression has been reported in several epilepsy patients [1,3]. However, the impact of neuron-astrocyte interactions on hyperexcitability on single cell level is underexplored. We developed a biophysical model of a presynaptic neuron and astrocyte to explore the relation between astrocytic EAAT2-mediated glutamate clearance, presynaptic glutamate receptors and bursting activity.MethodsWe build on our previous work [4,5], where we consider a presynaptic neuron and an astrocyte in a finite extracellular space (ECS). This model describes sodium, potassium, chloride dynamics as well as calcium-dependent exocytosis and glutamate-glutamine (GG) recycling. For this study, we add a potassium bath with diffusion to the ECS to induce neuronal bursting (Fig. 1A). Additionally, we implement the presynaptic glutamate receptors AMPA and NMDA, which are important in regulating hyperexcitability. Lastly, we study the impact of the antiseizure drugs perampanel and memantine by simulating the effect of these AMPA and NMDA receptor antagonists.ResultsWe induce neuronal bursting by increasing extracellular potassium in the bath. We first examine how AMPA and NMDA permeabilities affect burst frequency (Fig. 1C), where frequency refers to spike frequency during the last burst or during tonic firing, to fit the NMDA/AMPA ratio to experimental data [6]. Higher permeabilities increase neuronal firing and intracellular calcium, triggering a feedback loop that enhances neuronal glutamate release. Reducing EAAT permeability raises burst frequency and induces tonic firing (Fig. 1B). Finally, AMPA and NMDA antagonists, perampanel and memantine [7], reduce firing despite elevated extracellular glutamate, with perampanel showing a more significant reduction in firing frequency (Fig. 1D). DiscussionOur results show that reduced EAAT expression, as observed in several epilepsy patients, results in increased extracellular glutamate and overstimulation of excitatory glutamate receptors. Furthermore, we show that the AMPA and NMDA receptor permeabilities affect burst frequency. Receptor antagonists such as perampanel and memantine are able to reduce firing. In conclusion, our detailed neuron–astrocyte model provides insight into the effects of reduced EAAT expression and receptor antagonists on hyperexcitability.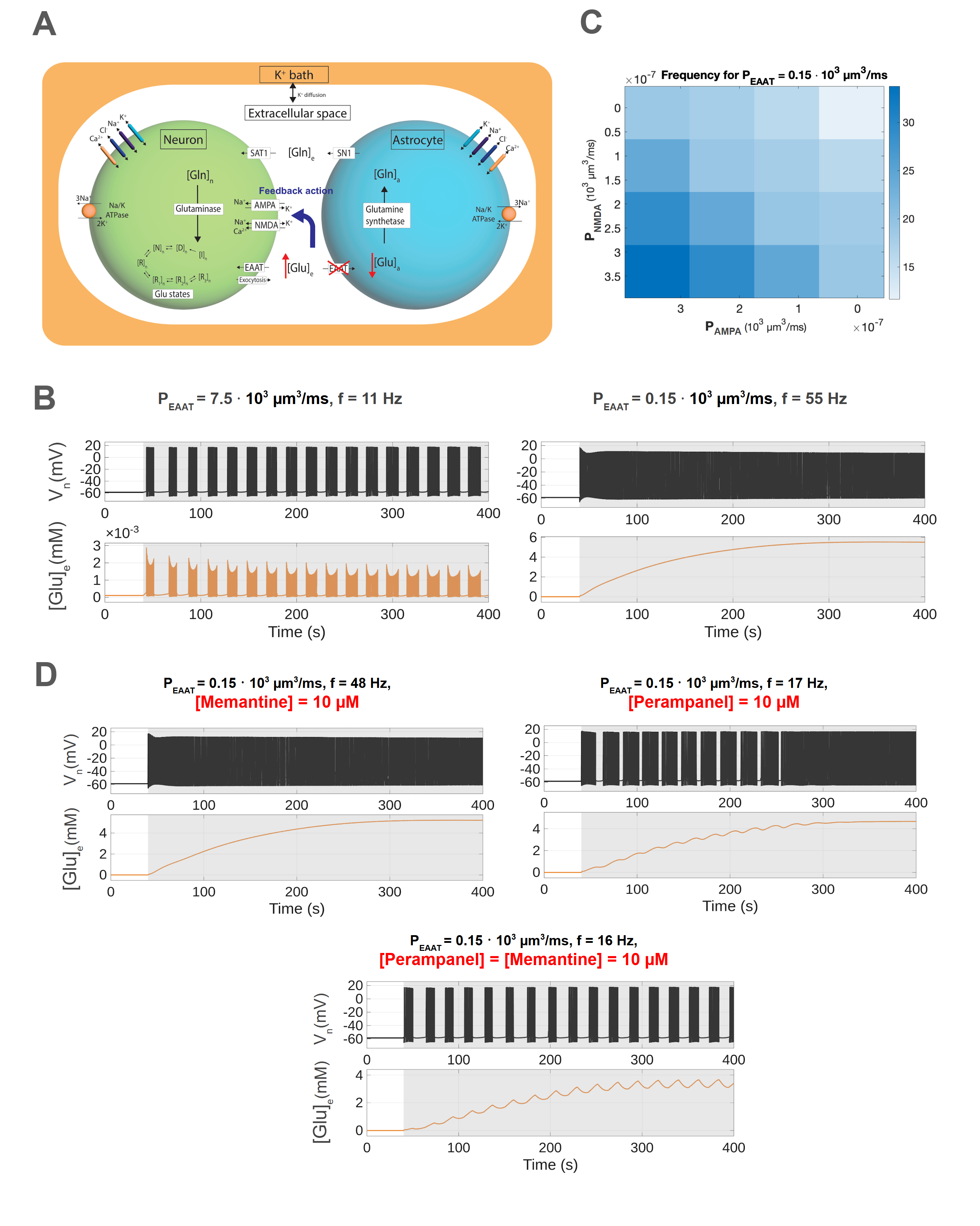 Figure 1. A: Three-compartment model illustrating the GG-cycle during EAAT2 knockout. B: The membrane potential, spike frequency f and ECS glutamate at different EAAT2 permeabilities. C: Spike frequency within bursts as a function of NMDA and AMPA receptor permeability. D: Neuronal activity at fixed EAAT2 permeability (PEAAT=0.15 * 103 µm3/ms) under antagonist conditions.References
Figure 1. A: Three-compartment model illustrating the GG-cycle during EAAT2 knockout. B: The membrane potential, spike frequency f and ECS glutamate at different EAAT2 permeabilities. C: Spike frequency within bursts as a function of NMDA and AMPA receptor permeability. D: Neuronal activity at fixed EAAT2 permeability (PEAAT=0.15 * 103 µm3/ms) under antagonist conditions.References[1] Green, J. L., dos Santos, W. F., & Fontana, A. C. K. (2021). Biochemical Pharmacology, 10.1016/j.bcp.2021.114786
[2] Scimemi, A., Tian, H., & Diamond, J. S. (2009). The Journal of Neuroscience, 10.1523/JNEUROSCI.4845-09.2009
[3] Barker-Haliski, M., & White, H. (2015). Cold Spring Harbor perspectives in medicine, 10.1101/cshperspect.a022863
[4] van Susteren, H., Rose, C. R., van Putten, M. J., & Meijer, H. G. (2025). bioRxiv, 10.1101/2025.11.10.687543
[5] Kalia, M., et al. (2021). PLOS Computational Biology, 10.1371/journal.pcbi.1009019
[6] Watt, A. J., Sjöström, P. J., Häusser, M., Nelson, S. B., & Turrigiano, G. G. (2004). Nature neuroscience, 10.1038/nn1220
[7] Chen, T.-S., Huang, T.-H., Lai, M.-C., & Huang, C.-W. (2023). Biomedicines, 10.3390/biomedicines11030783
AcknowledgmentsHVS, HGEM, MJAMVP funded from the DFG, FOR2795 ‘Synapses under stress’ to CRR (Prof. Dr. Christine R. Rose) (Ro2327/13-2 and 14-2).
GG, HS, and JH were supported by the ERDF-Project Brain dynamics, No. CZ.02.01.01/00/22\_008/0004643, a Lumina-Quaeruntur fellowship (LQ100302301), and the long-term strategic development financing of the Institute of Computer Science (RVO:67985807).
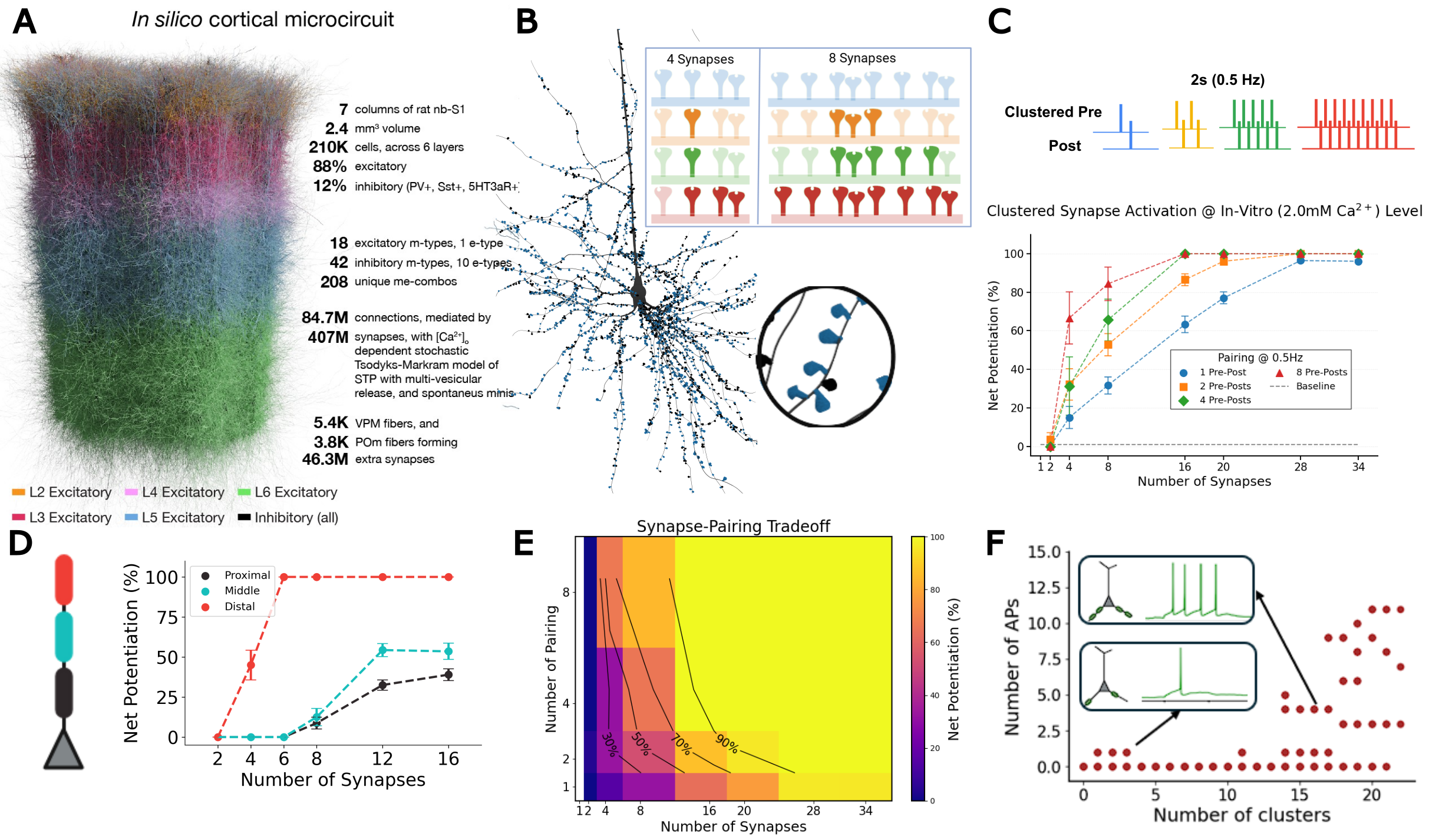 Figure 1. A, In silico cortical microcircuit. B, L5-TTPC with magnified cluster showing plasticity for 4 vs 8 co-active synapses. C, Clustered pre-post pairing (0.5 Hz); net potentiation vs synapse number. D, Spatial learning gradient E, Synapse-pairing tradeoff heatmap. F, Basal cluster coactivation triggers somatic burst.
Figure 1. A, In silico cortical microcircuit. B, L5-TTPC with magnified cluster showing plasticity for 4 vs 8 co-active synapses. C, Clustered pre-post pairing (0.5 Hz); net potentiation vs synapse number. D, Spatial learning gradient E, Synapse-pairing tradeoff heatmap. F, Basal cluster coactivation triggers somatic burst.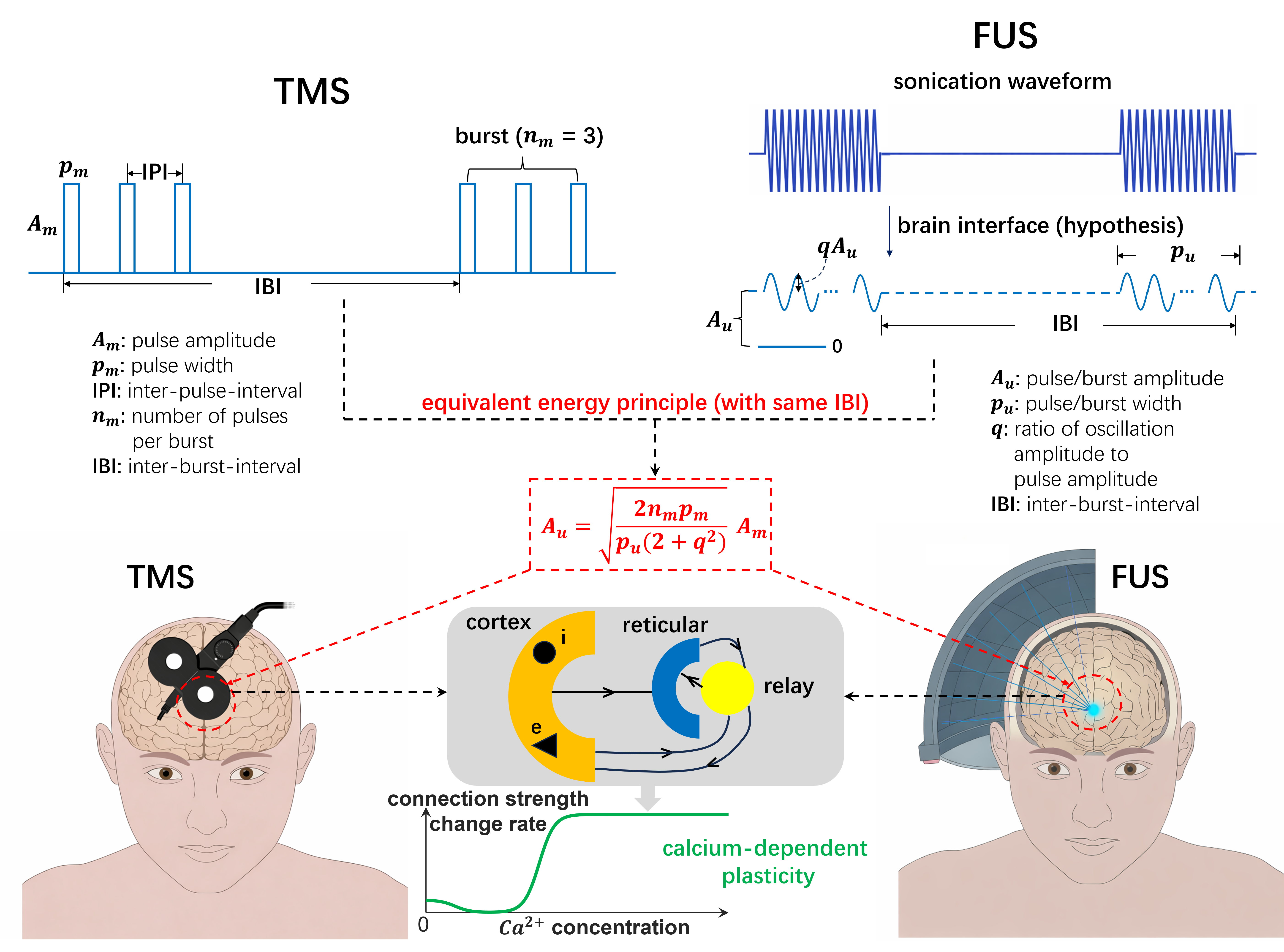 Figure 1. Equivalent energy principle for aligning rTMS and rTFUS plasticity models. rTMS delivers discrete pulse bursts; rTFUS delivers continuous bursts filtered by the skull interface to sub-1kHz sinusoids. The principle constrains parameters so both modalities deliver equal energy at the same carrier frequency, enabling unified mean-field modelling of calcium-dependent plasticity.
Figure 1. Equivalent energy principle for aligning rTMS and rTFUS plasticity models. rTMS delivers discrete pulse bursts; rTFUS delivers continuous bursts filtered by the skull interface to sub-1kHz sinusoids. The principle constrains parameters so both modalities deliver equal energy at the same carrier frequency, enabling unified mean-field modelling of calcium-dependent plasticity.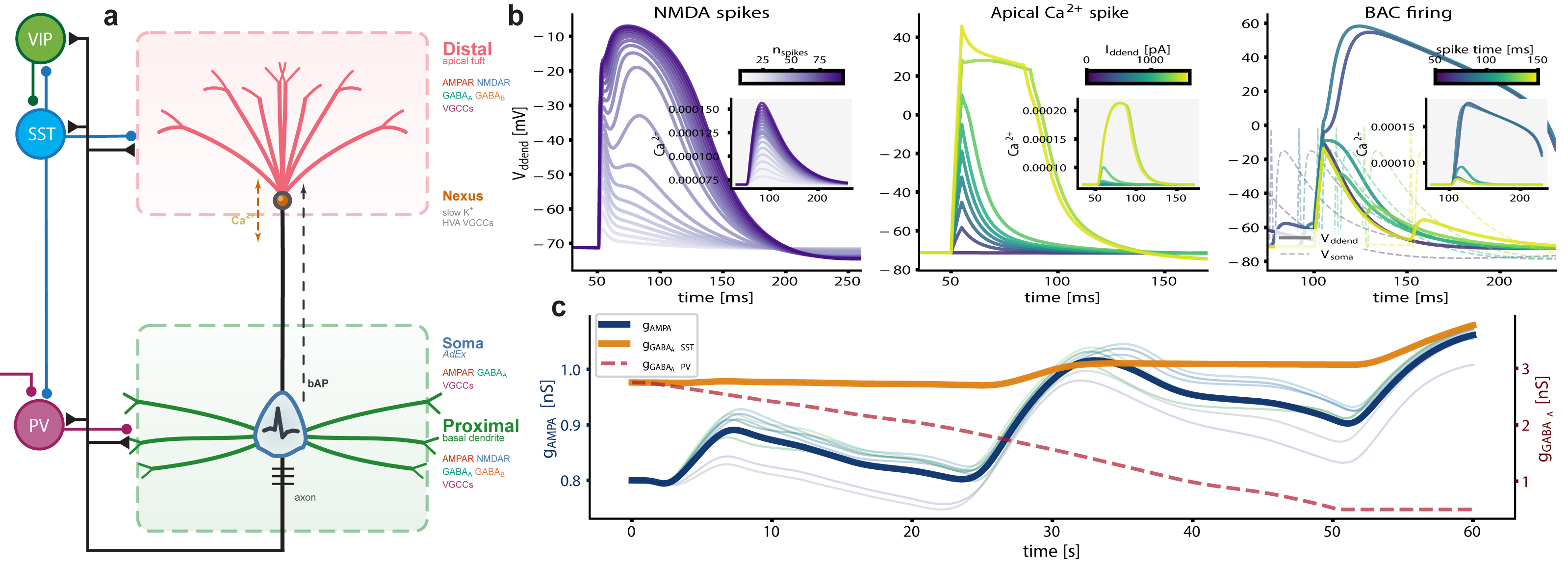 Figure 1. Augmented tripod neuron with compartment-specific electrogenic events and coupled E/I plasticity. (a) Circuit schematic with compartment-resolved receptors and interneuron targeting. (b) NMDA plateaus, apical Ca²⁺ spikes, and BAC firing with dendritic calcium transients (insets). (c) Shared calcium couples excitatory and inhibitory weight dynamics, actively maintaining E/I balance.References
Figure 1. Augmented tripod neuron with compartment-specific electrogenic events and coupled E/I plasticity. (a) Circuit schematic with compartment-resolved receptors and interneuron targeting. (b) NMDA plateaus, apical Ca²⁺ spikes, and BAC firing with dendritic calcium transients (insets). (c) Shared calcium couples excitatory and inhibitory weight dynamics, actively maintaining E/I balance.References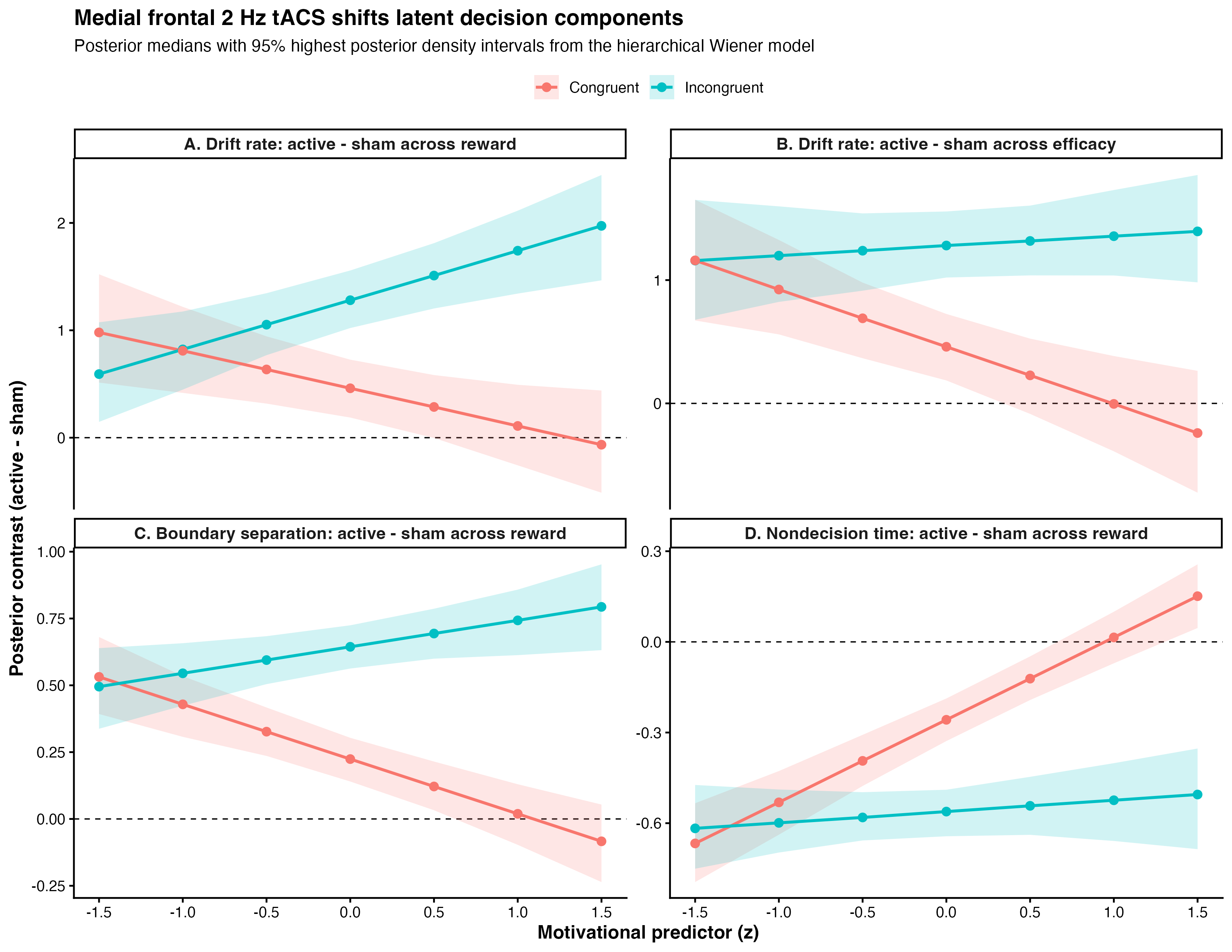 Figure 1. Active minus sham posterior contrasts from the hierarchical Wiener diffusion model. Top panels show drift-rate changes across reward and efficacy, separated by congruency. Bottom panels show boundary-separation and nondecision-time changes across reward. Points indicate posterior medians; ribbons show 95% highest posterior density intervals.
Figure 1. Active minus sham posterior contrasts from the hierarchical Wiener diffusion model. Top panels show drift-rate changes across reward and efficacy, separated by congruency. Bottom panels show boundary-separation and nondecision-time changes across reward. Points indicate posterior medians; ribbons show 95% highest posterior density intervals.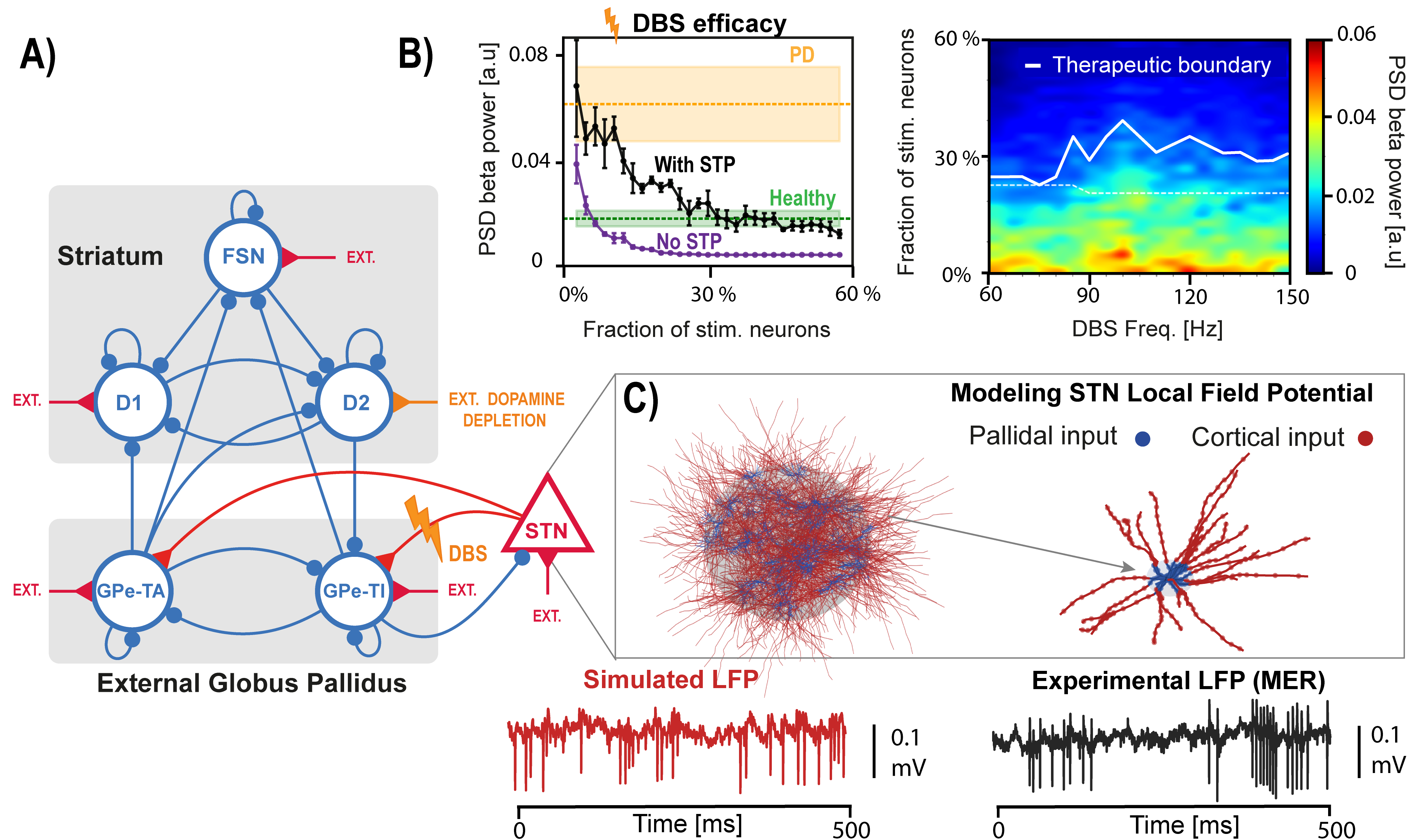 Figure 1. A) Spiking network model of the basal ganglia. B) Left: efficacy of STN DBS as a function of the fraction of stimulated neurons, with and without short-term plasticity (STP). Right: effect of stimulation frequencies on beta spectral power. C) Top: STN population and morphological neuron model receiving cortical and pallidal inputs. Bottom: simulated and recorded local field potentials (LFPs).
Figure 1. A) Spiking network model of the basal ganglia. B) Left: efficacy of STN DBS as a function of the fraction of stimulated neurons, with and without short-term plasticity (STP). Right: effect of stimulation frequencies on beta spectral power. C) Top: STN population and morphological neuron model receiving cortical and pallidal inputs. Bottom: simulated and recorded local field potentials (LFPs).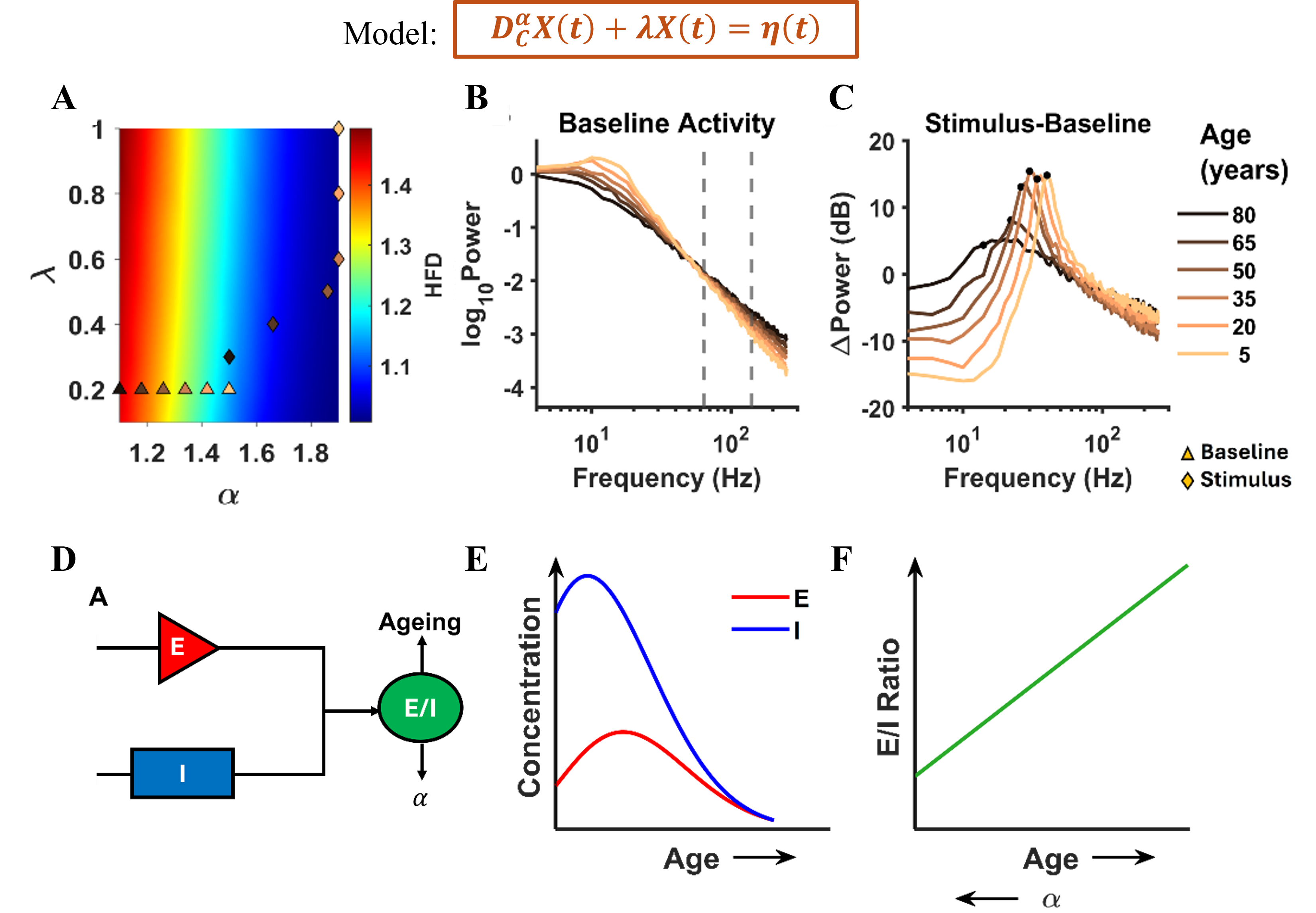 Figure 1. Age-related inferences: Top: Model equation. (A) HFD variation with α and λ. (B), (C) The PSDs and ΔPower (in dB) corresponding to triangles and diamonds respectively. (D) Illustration of E/I dependence on ageing and α. (E) The concentration of excitatory and inhibitory neurotransmitters varying non-monotonically with age. (F) Corresponding monotonic E/I ratio with age and α.
Figure 1. Age-related inferences: Top: Model equation. (A) HFD variation with α and λ. (B), (C) The PSDs and ΔPower (in dB) corresponding to triangles and diamonds respectively. (D) Illustration of E/I dependence on ageing and α. (E) The concentration of excitatory and inhibitory neurotransmitters varying non-monotonically with age. (F) Corresponding monotonic E/I ratio with age and α.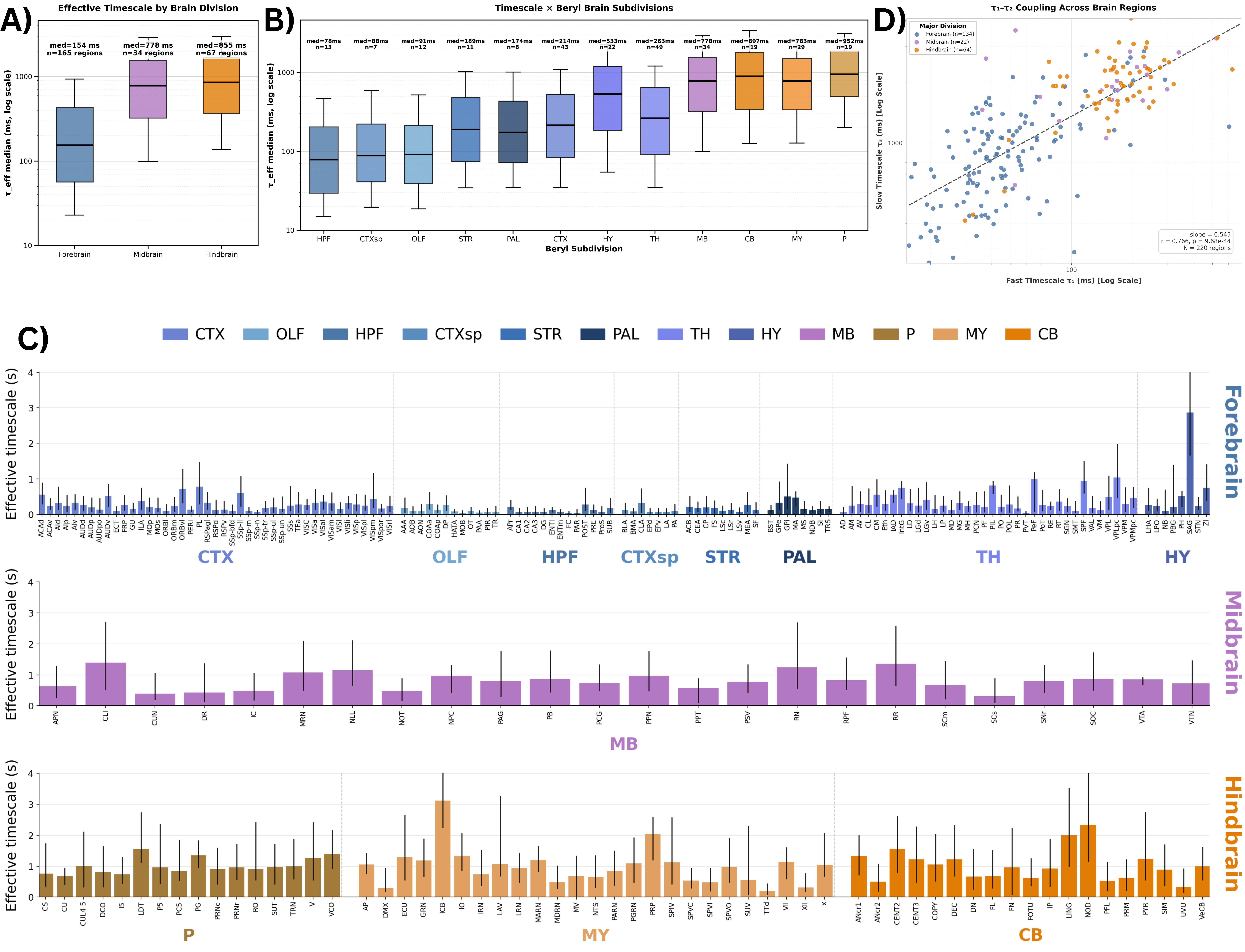 Brain-wide map of intrinsic neural timescales. (A) τ_eff by major brain division. (B) τ_eff across 12 brain subdivisions. (C) Median τ_eff per region; error bars show 10th–90th percentiles. Regions grouped by division, ordered alphabetically; colors denote subdivision. (D) Fast (τ₁) vs slow (τ₂) timescales across regions.
Brain-wide map of intrinsic neural timescales. (A) τ_eff by major brain division. (B) τ_eff across 12 brain subdivisions. (C) Median τ_eff per region; error bars show 10th–90th percentiles. Regions grouped by division, ordered alphabetically; colors denote subdivision. (D) Fast (τ₁) vs slow (τ₂) timescales across regions.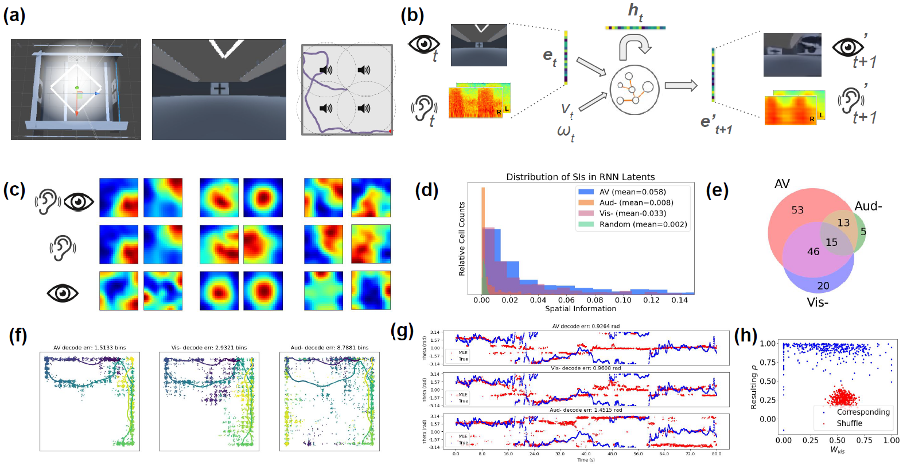 (a) Virtual environment with visual cues and sound sources; (b) Model architecture and pipeline; (c) Spatial ratemap examples in audiovisual, and lesioned (-) conditions; (d) Distribution of spatial information contents; (e) Number of place units identified; (f) ML decoding of position; (g) ML decoding of head direction; (h) Distribution of visual weights (x) and resulting correlations (y); compar
(a) Virtual environment with visual cues and sound sources; (b) Model architecture and pipeline; (c) Spatial ratemap examples in audiovisual, and lesioned (-) conditions; (d) Distribution of spatial information contents; (e) Number of place units identified; (f) ML decoding of position; (g) ML decoding of head direction; (h) Distribution of visual weights (x) and resulting correlations (y); compar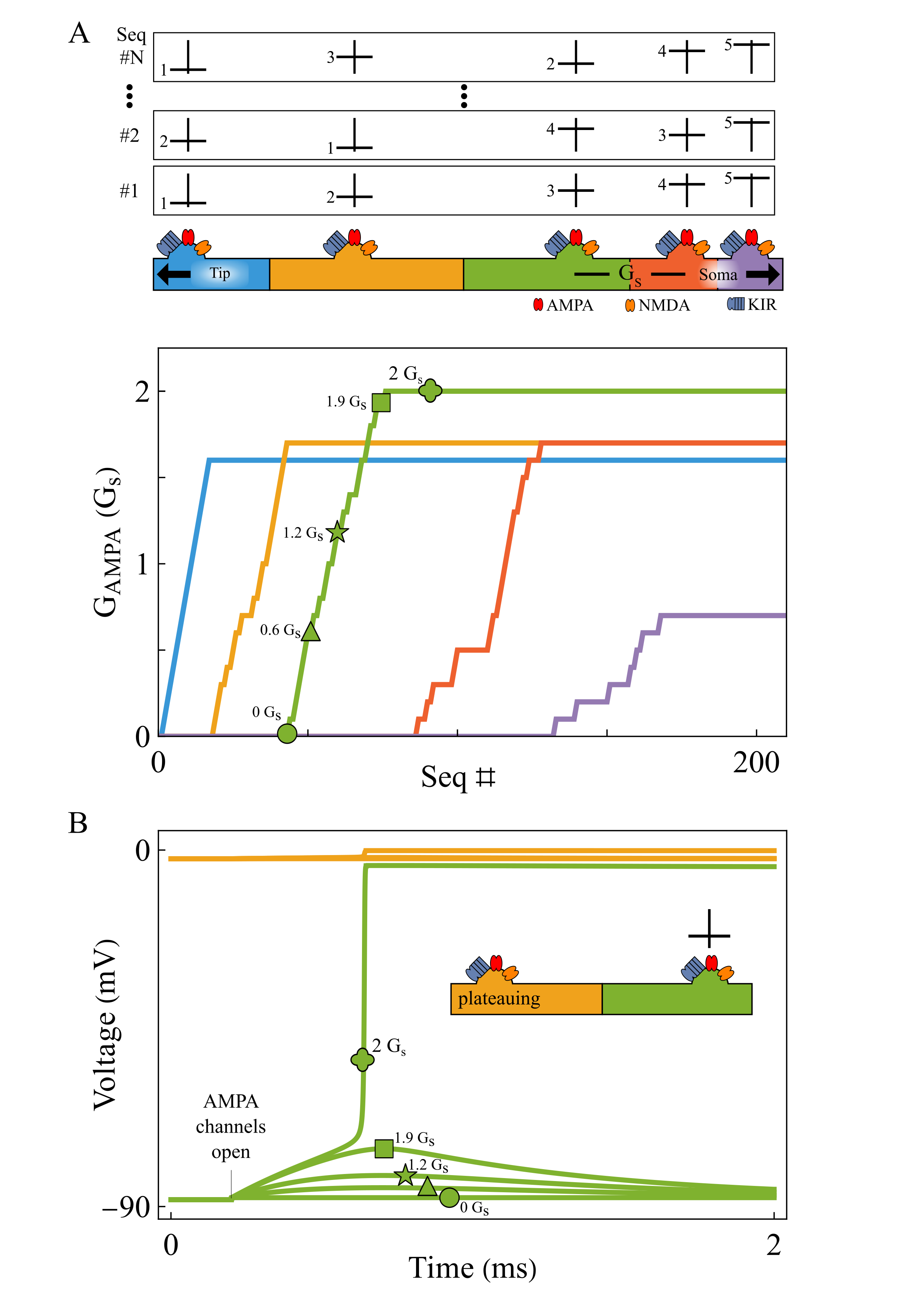 Local AMPA plasticity tunes dendritic sequence selectivity. Repeated tip-to-soma and shuffled inputs progressively strengthen AMPA conductance from tip to soma (A). When a segment (green) receives input while its tip-side neighbor is plateauing (yellow), weak AMPA initially fails to trigger a plateau. As AMPA increases, the segment abruptly transitions to plateau at a bifurcation (B).References
Local AMPA plasticity tunes dendritic sequence selectivity. Repeated tip-to-soma and shuffled inputs progressively strengthen AMPA conductance from tip to soma (A). When a segment (green) receives input while its tip-side neighbor is plateauing (yellow), weak AMPA initially fails to trigger a plateau. As AMPA increases, the segment abruptly transitions to plateau at a bifurcation (B).References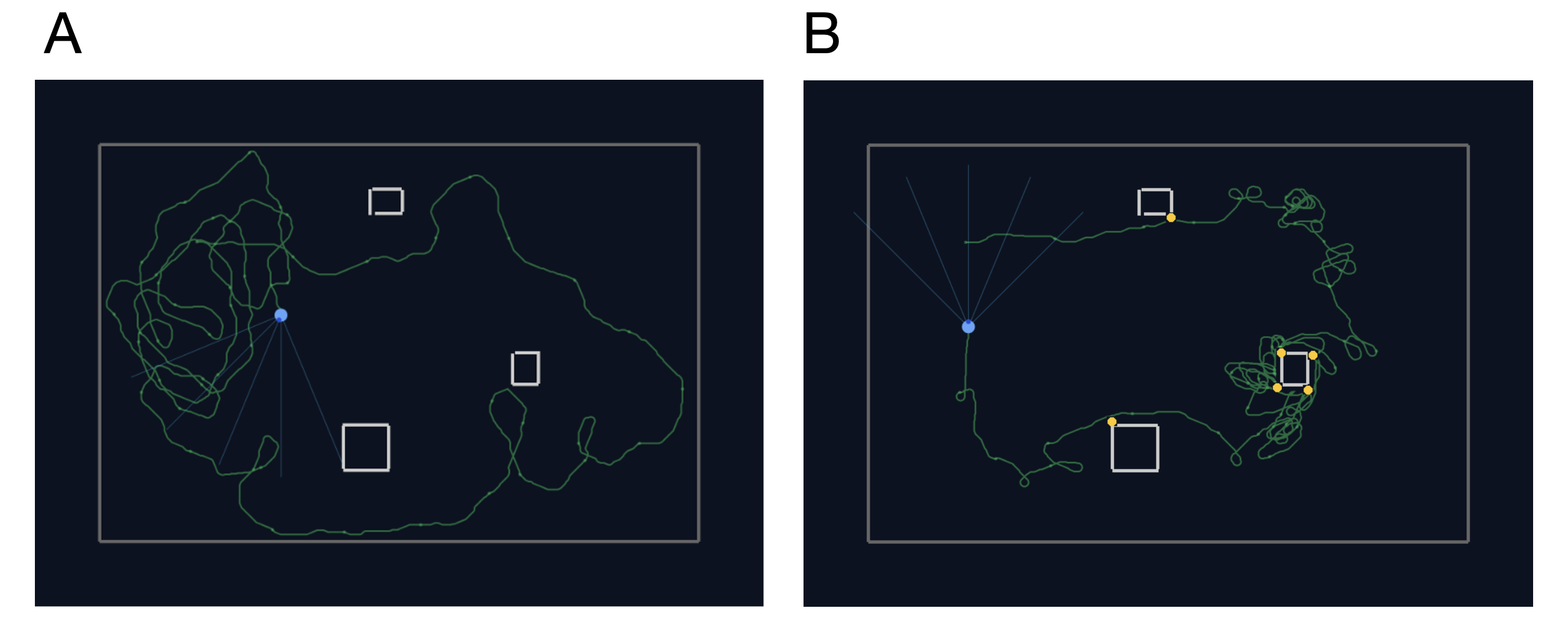 Example trajectories (green) of an agent (light blue circle with blue rays indicating visual sensors) in an arena with three high-contrast obstacles. A: The agent forages where no obstacles are present. It also frequents the part of the arena with obstacles, but never collides with them. B: Trajectory for an agent with less training times, leading to frequent collisions (yellow dots).References
Example trajectories (green) of an agent (light blue circle with blue rays indicating visual sensors) in an arena with three high-contrast obstacles. A: The agent forages where no obstacles are present. It also frequents the part of the arena with obstacles, but never collides with them. B: Trajectory for an agent with less training times, leading to frequent collisions (yellow dots).References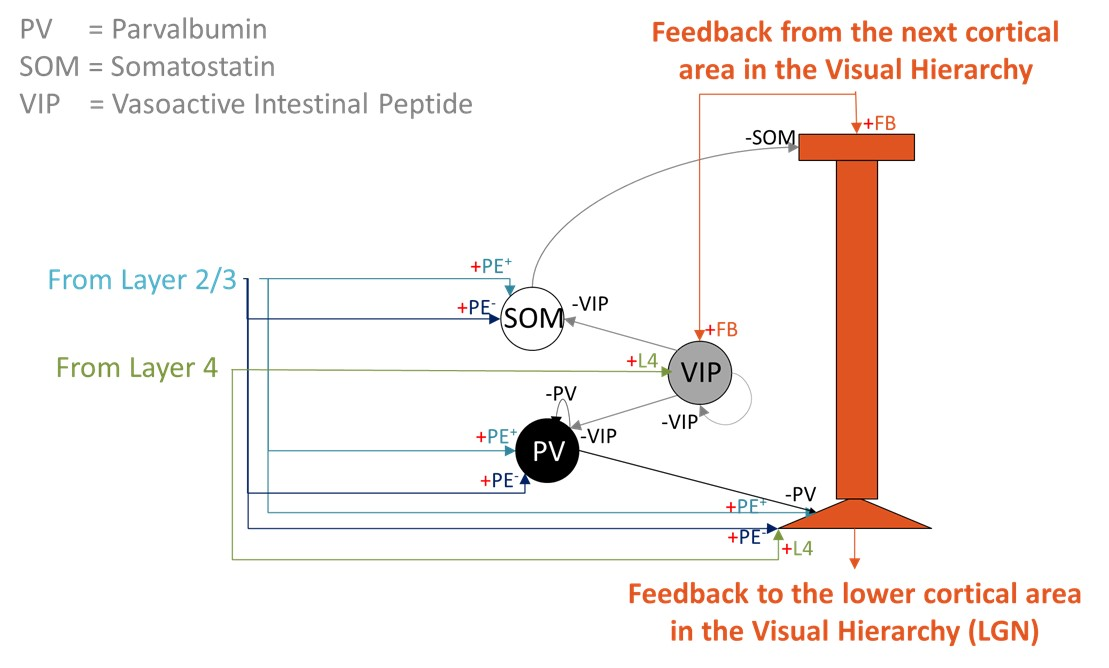 Layer 5 predictive-coding microcircuit. A two-compartment Layer 5 pyramidal neuron integrates somatic input from Layer 2/3 prediction-error populations (PE+ and PE-) and Layer 4 feature neurons with apical feedback from higher cortical areas. A local VIP-PV-SOM motif regulates apical Ca2+ spikes and burst output.
Layer 5 predictive-coding microcircuit. A two-compartment Layer 5 pyramidal neuron integrates somatic input from Layer 2/3 prediction-error populations (PE+ and PE-) and Layer 4 feature neurons with apical feedback from higher cortical areas. A local VIP-PV-SOM motif regulates apical Ca2+ spikes and burst output.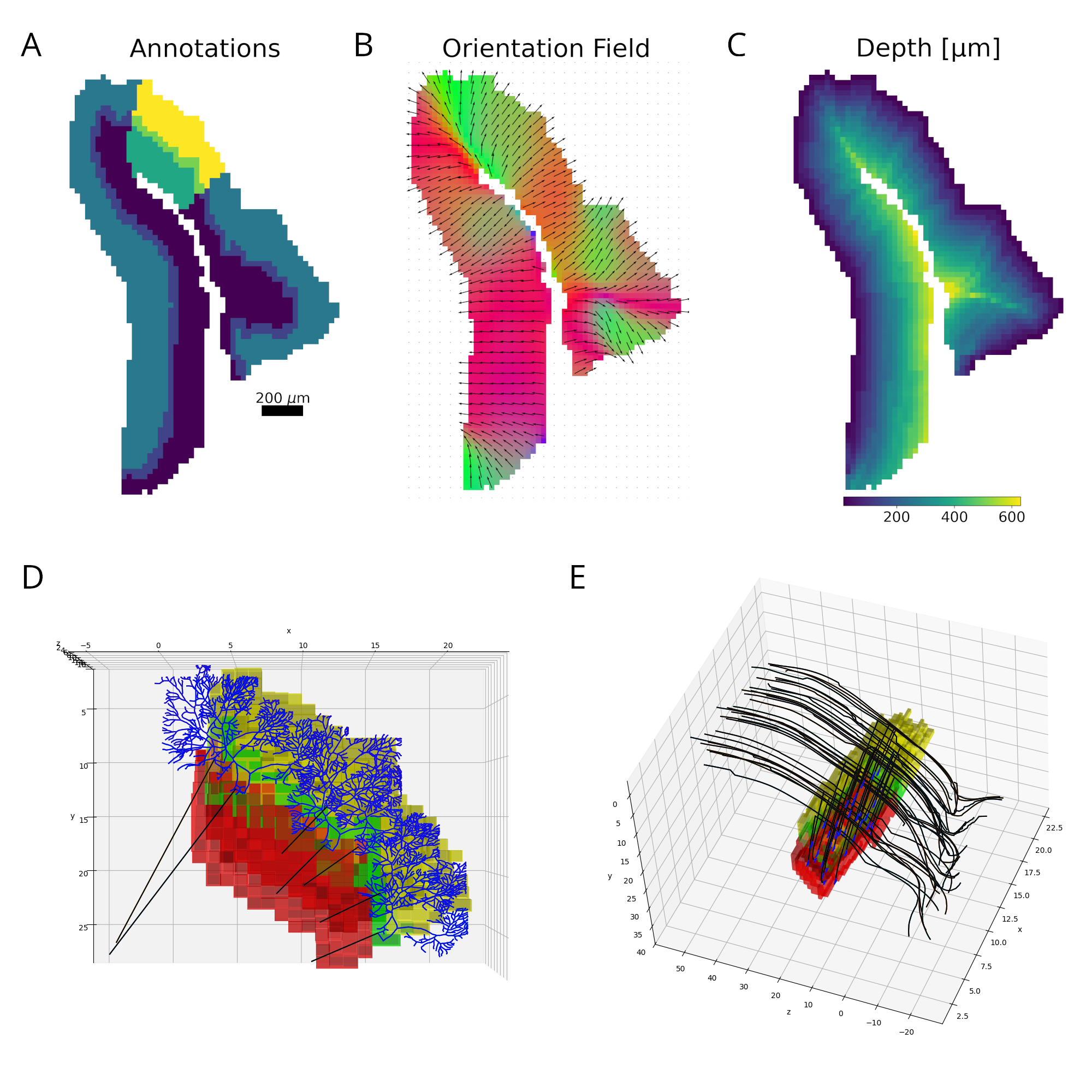 Figure 1. Reconstruction pipeline. A. Declive layers shown in colors with the selected column highlighted. B. Orientation field showing the local axons’ main axis. Colors represent the vectors’ norm. C. Distance to the outside border, following the orientation field. D. E. Purkinje and granule cells´ morphology scaled and bent according to the declive shape.
Figure 1. Reconstruction pipeline. A. Declive layers shown in colors with the selected column highlighted. B. Orientation field showing the local axons’ main axis. Colors represent the vectors’ norm. C. Distance to the outside border, following the orientation field. D. E. Purkinje and granule cells´ morphology scaled and bent according to the declive shape.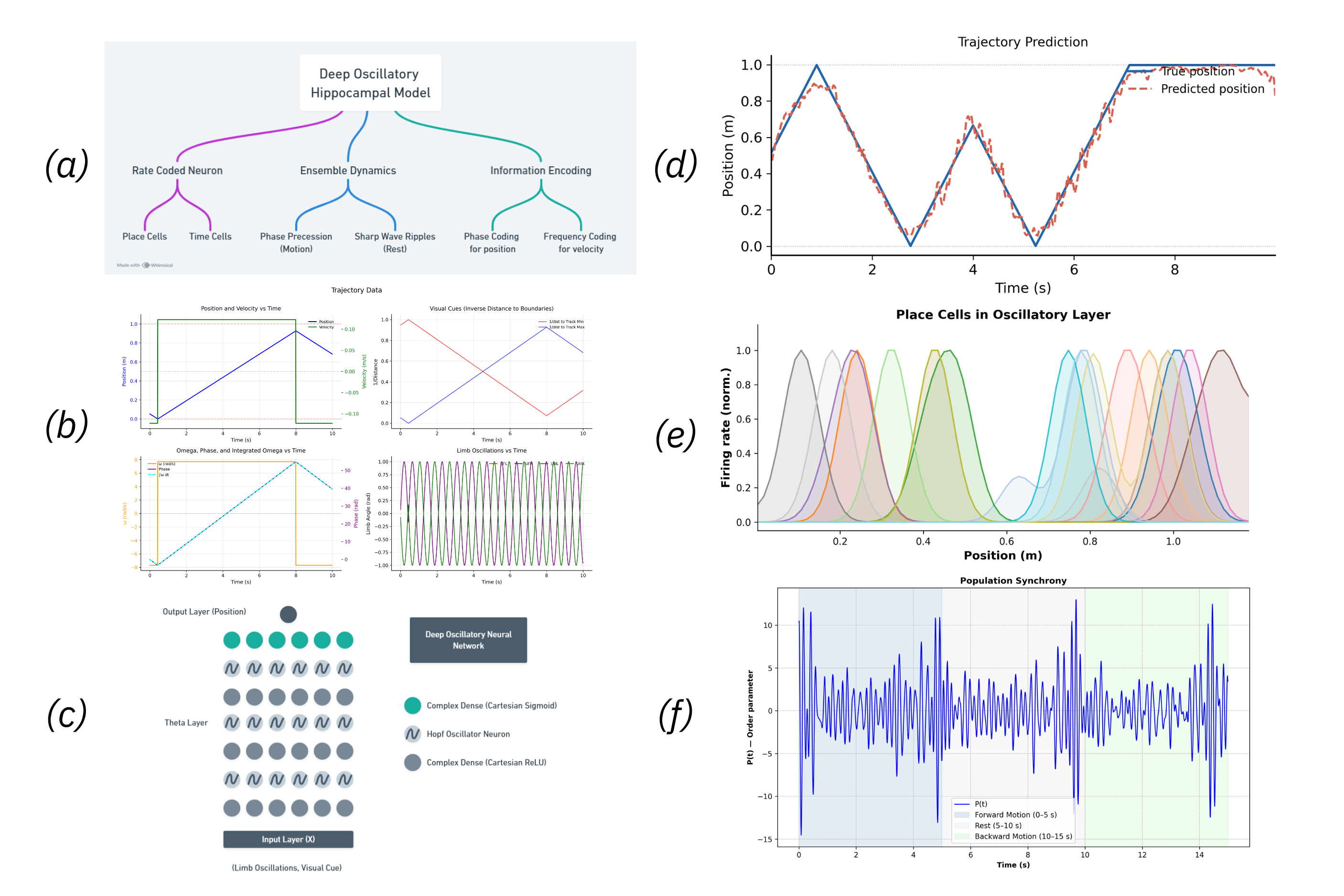 Figure 1. (a) Model Flowchart, (b) Input Data, (c) Oscillatory Neural Network Diagram, (d) Trajectory Prediction, (e) Place Cells - different colors correspond to different neurons, (f) Sharp Wave Ripples
Figure 1. (a) Model Flowchart, (b) Input Data, (c) Oscillatory Neural Network Diagram, (d) Trajectory Prediction, (e) Place Cells - different colors correspond to different neurons, (f) Sharp Wave Ripples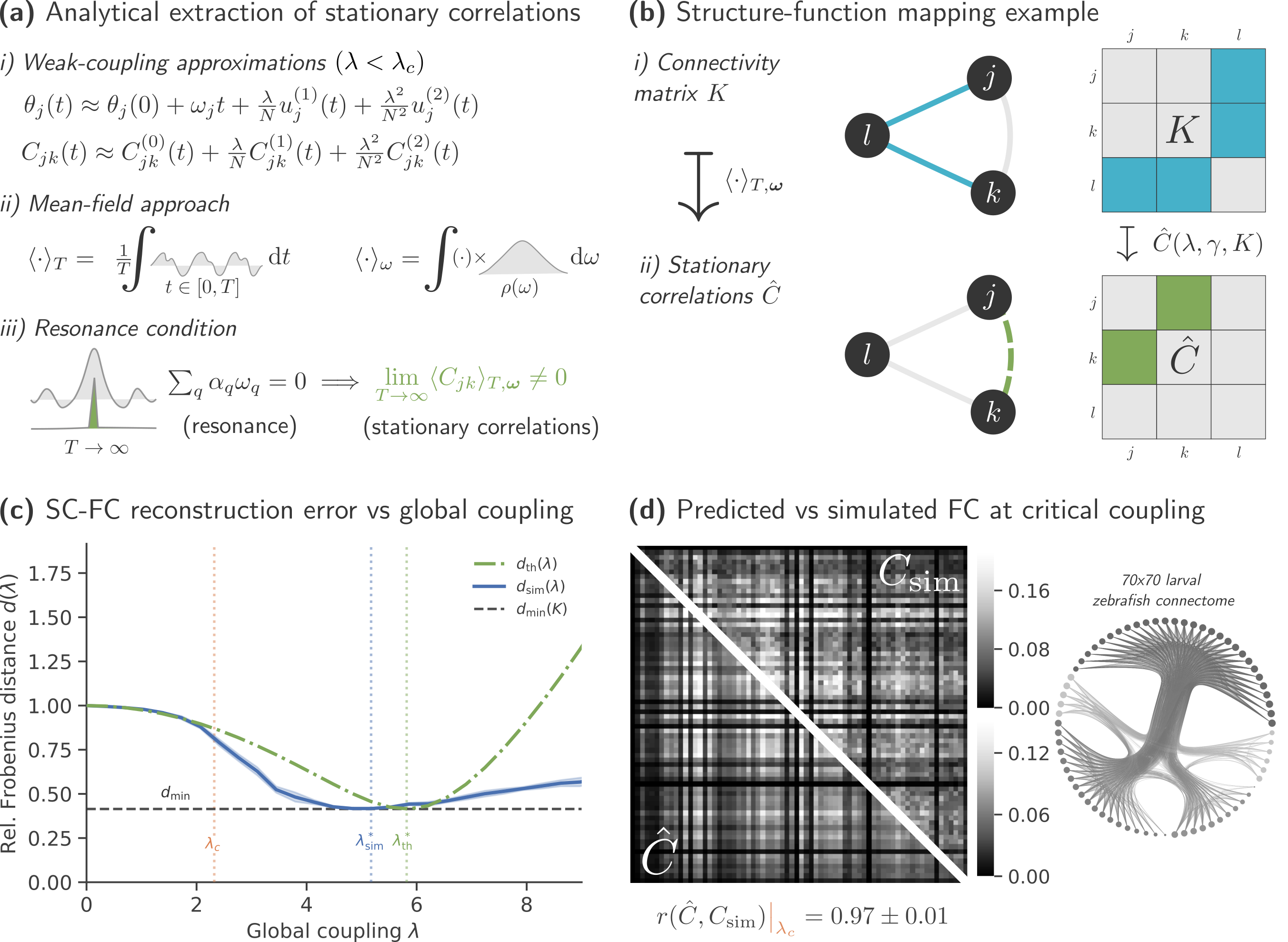 Figure 1. Predicting coactivity from anatomy in neural oscillators. (a) Derivation of predicted functional connectivity: phase trajectories are expanded, correlations averaged, and stationary terms selected. (b) Example for N=2 oscillators. (c) SC–FC reconstruction error follows theory up to synchronization (λc = 2.32). (d) Predicted and simulated FC remain highly similar (cosine similarity ≥ 0.97).
Figure 1. Predicting coactivity from anatomy in neural oscillators. (a) Derivation of predicted functional connectivity: phase trajectories are expanded, correlations averaged, and stationary terms selected. (b) Example for N=2 oscillators. (c) SC–FC reconstruction error follows theory up to synchronization (λc = 2.32). (d) Predicted and simulated FC remain highly similar (cosine similarity ≥ 0.97).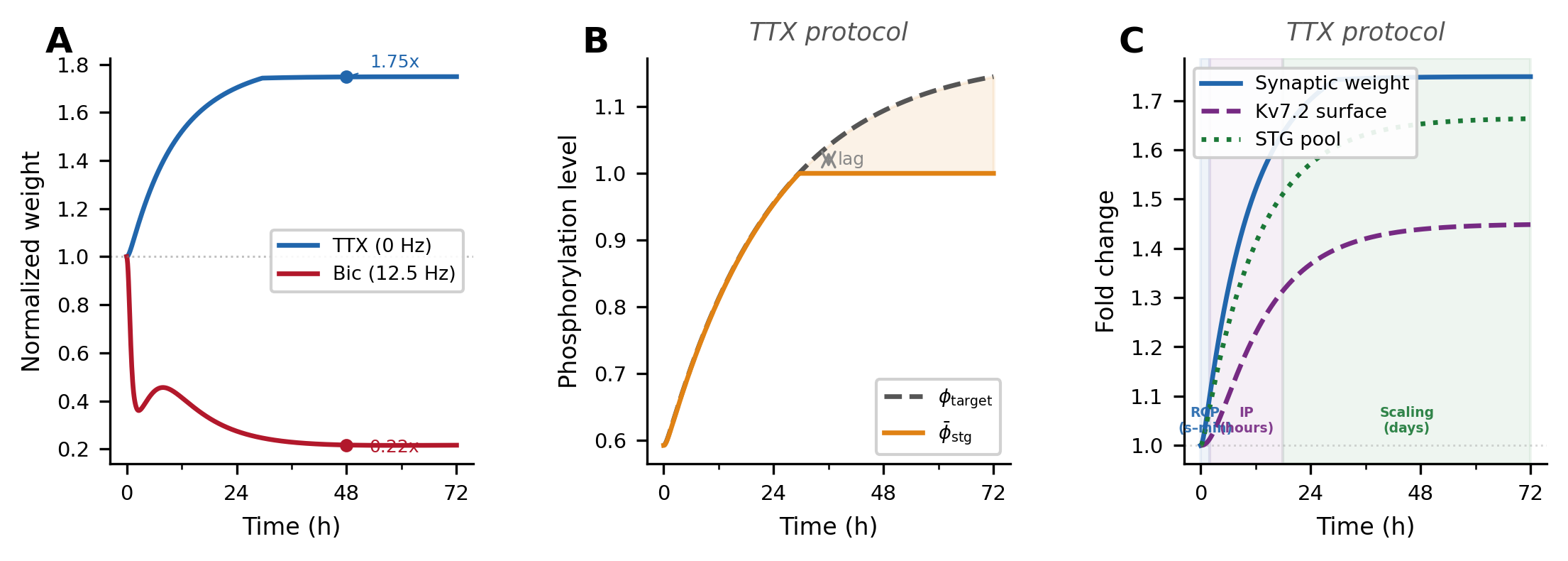 Figure 1. Simulated homeostatic response to inactivity (TTX). (A) Multiplicative synaptic scaling: upscaling (1.75×) and downscaling (0.22×) at 48 h. (B) Stargazin phosphorylation (φ̄_stg) lags the homeostatic target (φ_target) due to enzymatic inertia. (C) Three compensatory tiers emerge sequentially: AMPAR redistribution (seconds–min), Kv7.2 adjustment (hours), and synaptic scaling (days).
Figure 1. Simulated homeostatic response to inactivity (TTX). (A) Multiplicative synaptic scaling: upscaling (1.75×) and downscaling (0.22×) at 48 h. (B) Stargazin phosphorylation (φ̄_stg) lags the homeostatic target (φ_target) due to enzymatic inertia. (C) Three compensatory tiers emerge sequentially: AMPAR redistribution (seconds–min), Kv7.2 adjustment (hours), and synaptic scaling (days).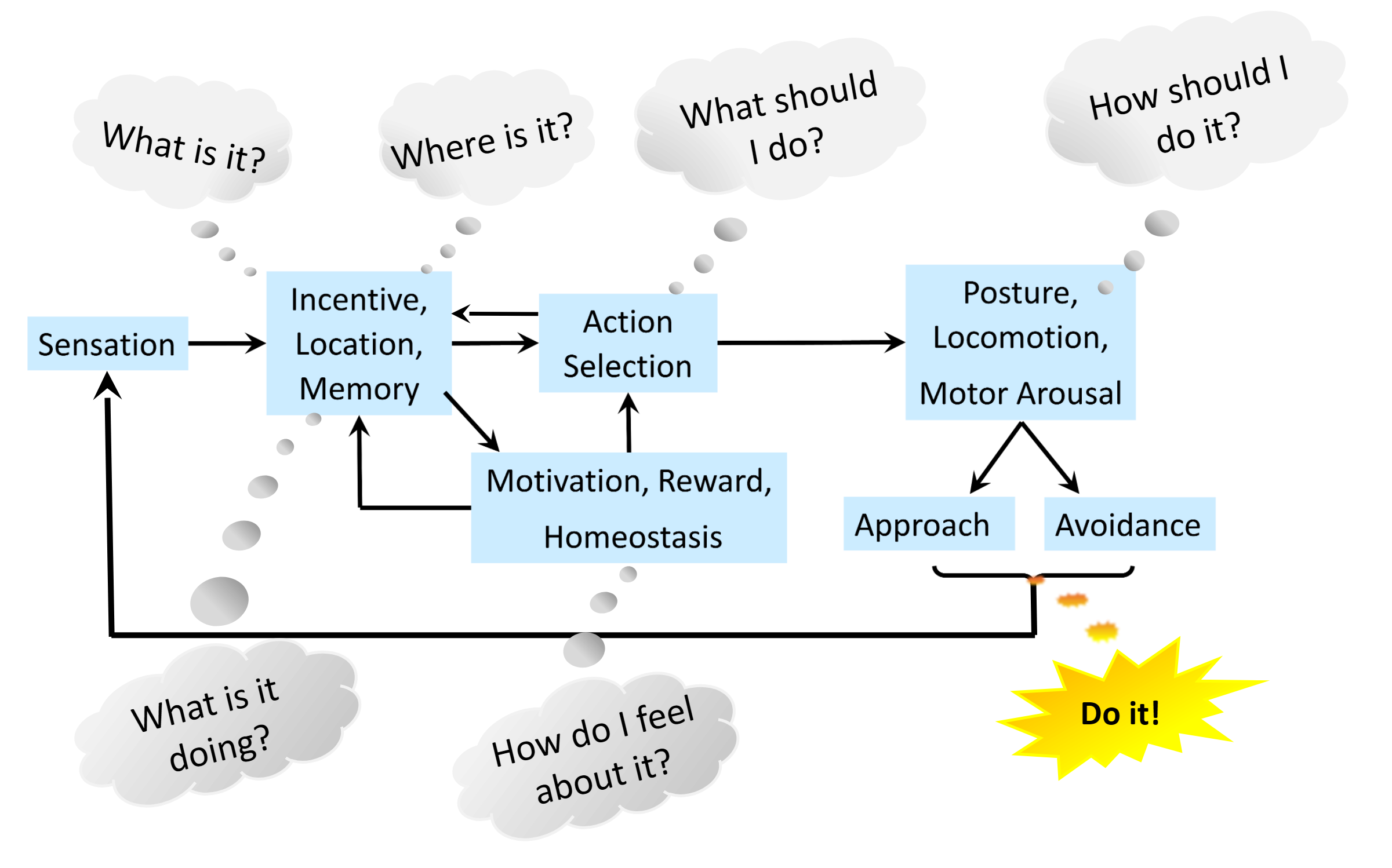 Figure 1. Both simple and complex animals handle flow of information from sensation to behavior with a common modular nervous system organization. Stimuli characteristics, incentives and locations are integrated with memory, motivation and affect for decisive action selection, with 5 critical computations from “What is it?” to “How should I do it?”, and cognitive mapping adding “What is it doing?”.
Figure 1. Both simple and complex animals handle flow of information from sensation to behavior with a common modular nervous system organization. Stimuli characteristics, incentives and locations are integrated with memory, motivation and affect for decisive action selection, with 5 critical computations from “What is it?” to “How should I do it?”, and cognitive mapping adding “What is it doing?”.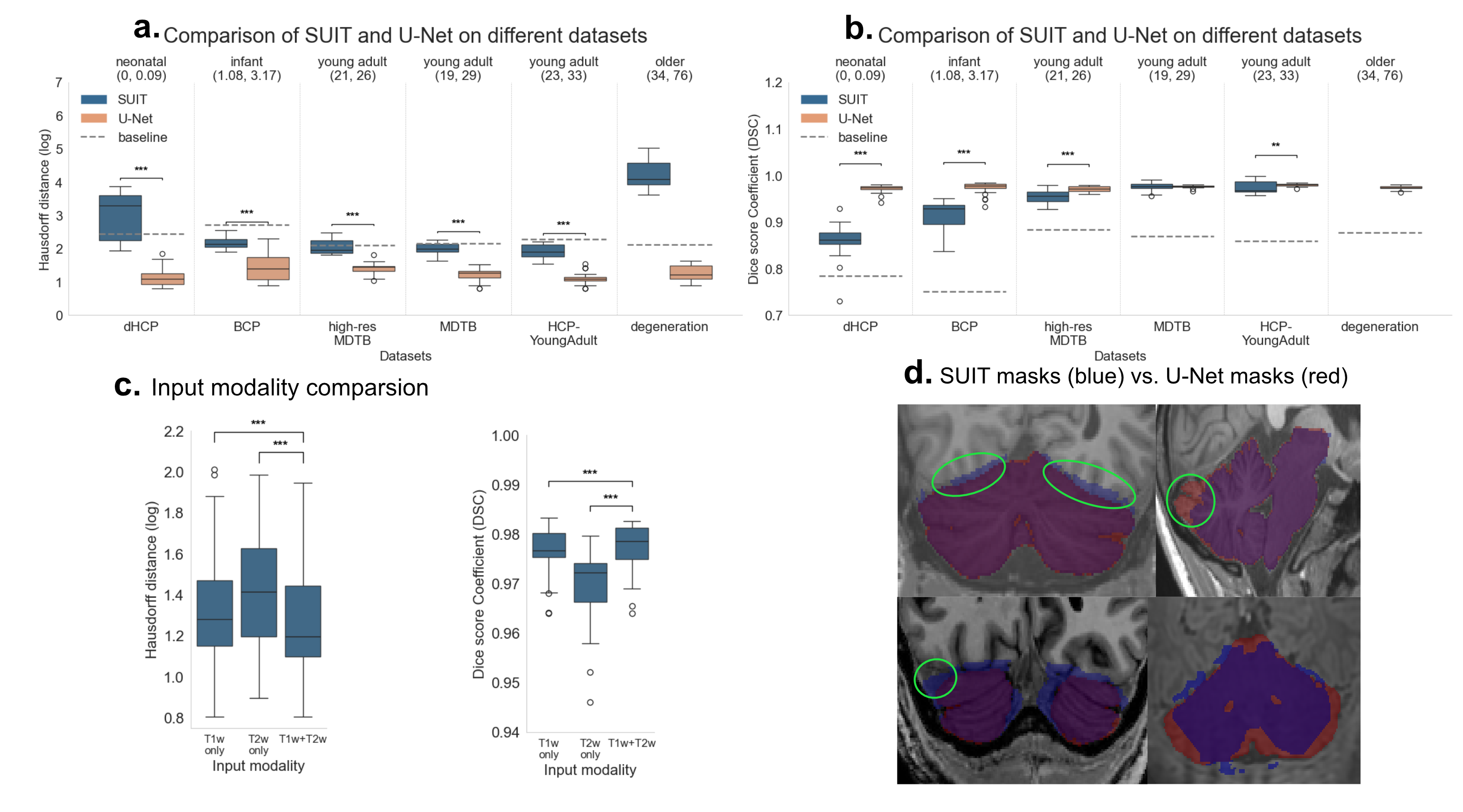 Figure 1. Isolation analysis. Comparison of U-Net VS SUIT for (a) Hausdorff Distance (HD) and (b) Dice Score Coefficient (DSC). c shows performance for different input modalities in the full datasets. Horizontal lines between bars with asterisks denote significant differences (paired t-tests). Baseline: Average mask prediction. d shows resulting mask in problematic subjects.
Figure 1. Isolation analysis. Comparison of U-Net VS SUIT for (a) Hausdorff Distance (HD) and (b) Dice Score Coefficient (DSC). c shows performance for different input modalities in the full datasets. Horizontal lines between bars with asterisks denote significant differences (paired t-tests). Baseline: Average mask prediction. d shows resulting mask in problematic subjects.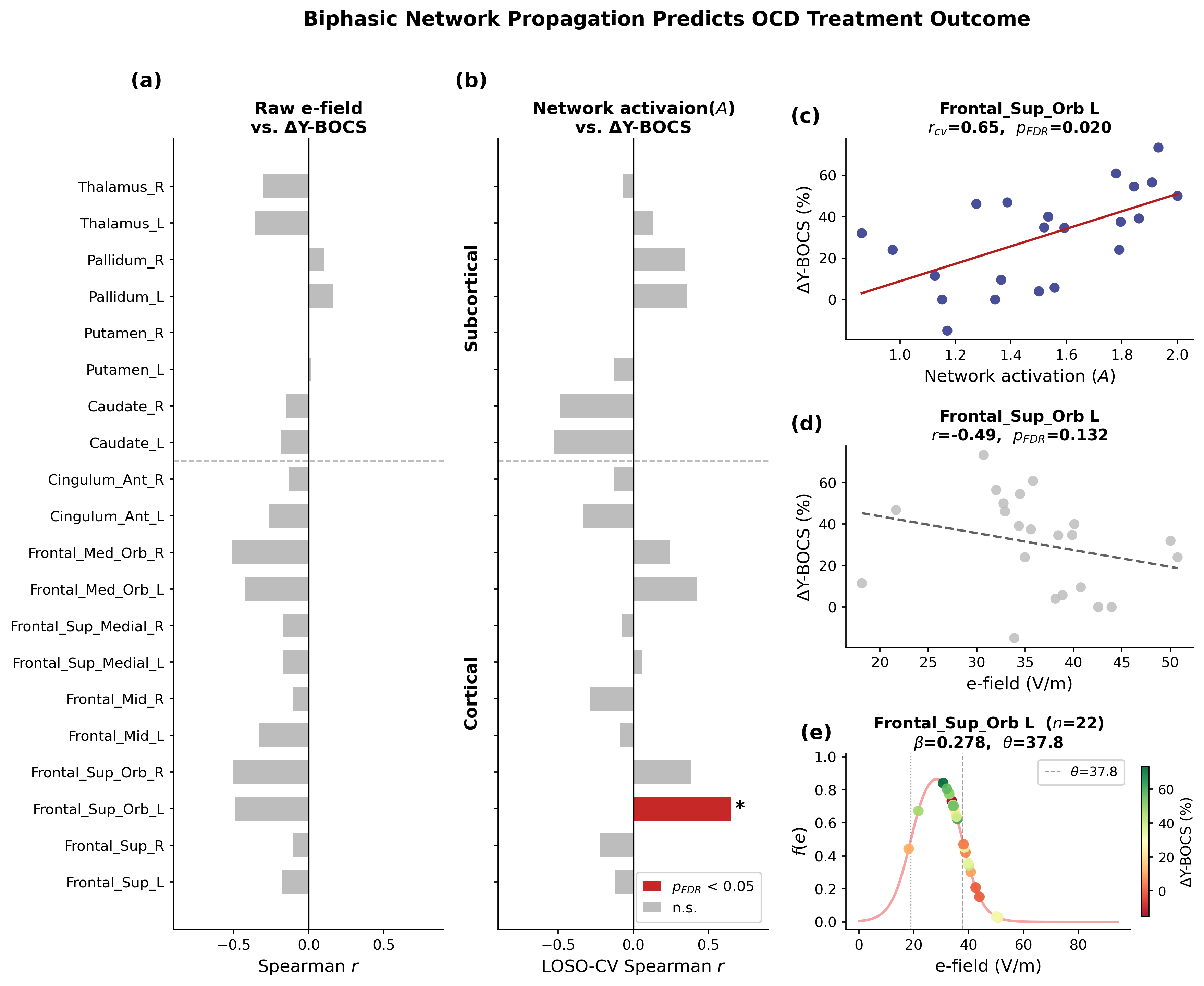 Figure 1. Correlations between E-field/network activation and clinical outcome in the CSTC ROIs. (a) Raw E-field showed no significant correlations. (b) The biphasic neuronal response model identified the left superior orbital frontal cortex as significantly associated with percentage Y-BOCS improvement. (c-f) Network activation/raw E-field correlation maps, and fitted biphasic neuronal response fReferences
Figure 1. Correlations between E-field/network activation and clinical outcome in the CSTC ROIs. (a) Raw E-field showed no significant correlations. (b) The biphasic neuronal response model identified the left superior orbital frontal cortex as significantly associated with percentage Y-BOCS improvement. (c-f) Network activation/raw E-field correlation maps, and fitted biphasic neuronal response fReferences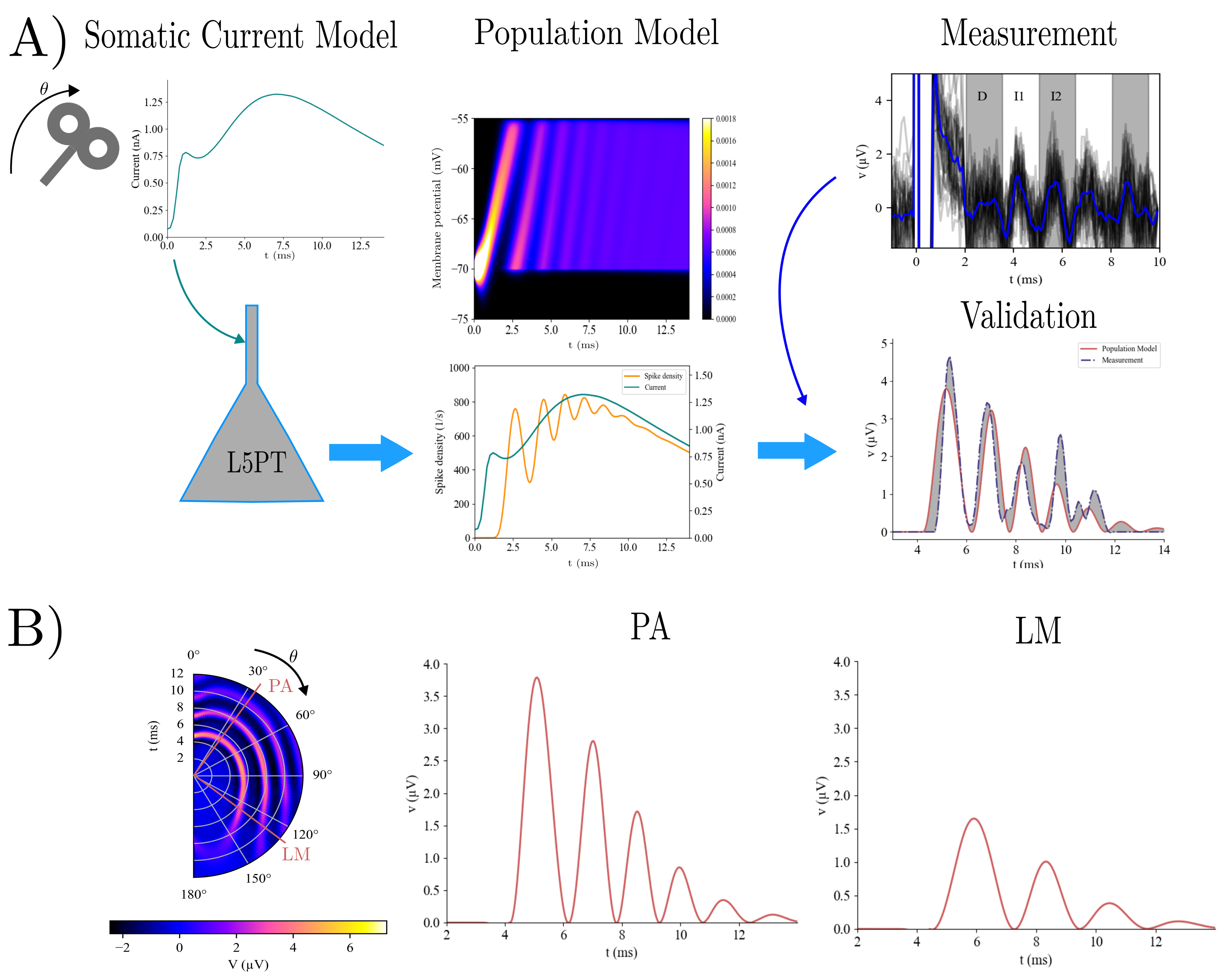 Figure 1. A) The somatic current model [3] generates coil orientation-sensitive somatic currents (first column). They are then applied to the L5PT model which computes membrane potential distribution and spike density for it (second column). This is then transformed to a potential and compared to measured I-waves. B) Orientation dependency of I-waves (first column) and potentials for putative parietal-anter
Figure 1. A) The somatic current model [3] generates coil orientation-sensitive somatic currents (first column). They are then applied to the L5PT model which computes membrane potential distribution and spike density for it (second column). This is then transformed to a potential and compared to measured I-waves. B) Orientation dependency of I-waves (first column) and potentials for putative parietal-anter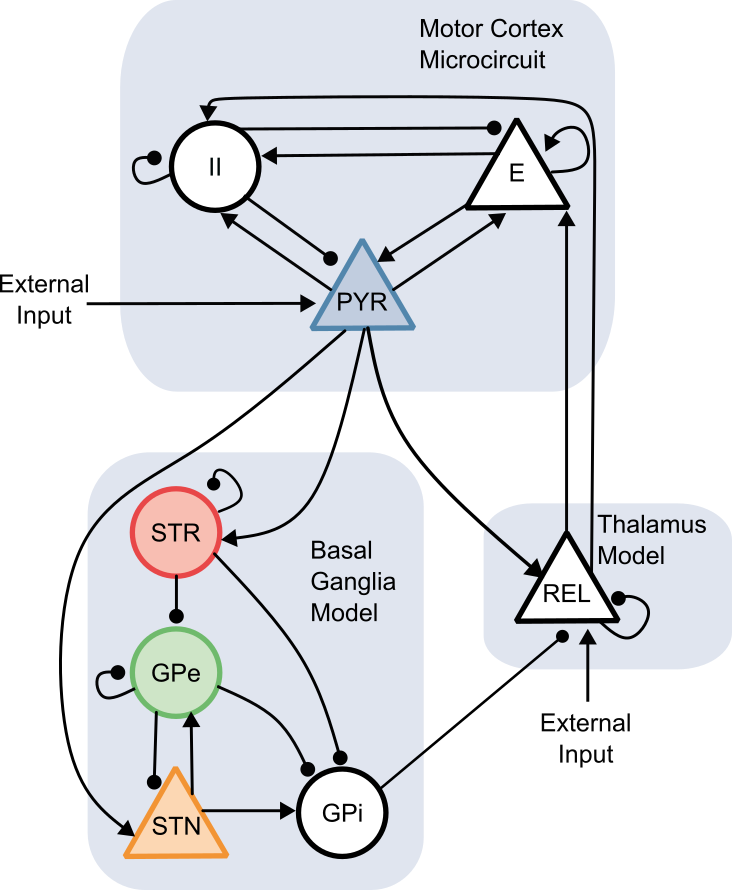 Figure 1. The BGTC circuit connectivity implemented in the neural mass model includes cortical excitatory (E), inhibitory interneuron (II), and deep pyramidal (DP) populations, as well as the striatum, globus pallidus externus (GPe), globus pallidus internus (GPi), subthalamic nucleus (STN), and thalamic relay nuclei (REL). Shaded regions indicate nodes with spectral data used for model fitting.
Figure 1. The BGTC circuit connectivity implemented in the neural mass model includes cortical excitatory (E), inhibitory interneuron (II), and deep pyramidal (DP) populations, as well as the striatum, globus pallidus externus (GPe), globus pallidus internus (GPi), subthalamic nucleus (STN), and thalamic relay nuclei (REL). Shaded regions indicate nodes with spectral data used for model fitting.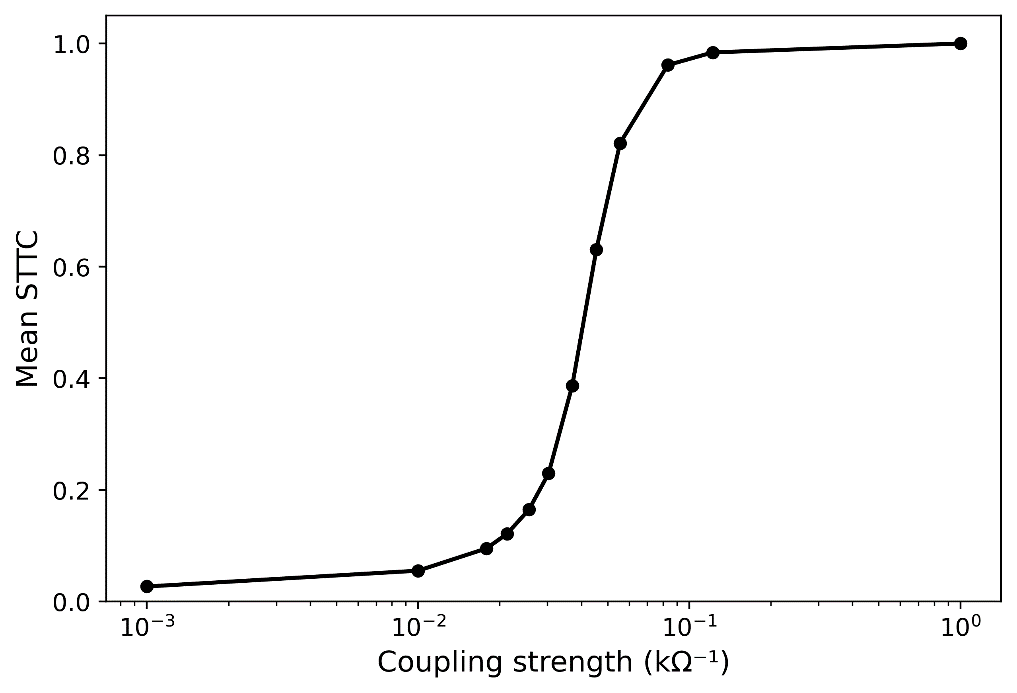 Figure 1. Mean spike time tiling coefficient (STTC) as a function of coupling strength in a network of 36 coupled stochastic relaxation-type oscillators. Increasing coupling drives the system from weakly correlated spiking activity toward global synchronization, indicating a continuous synchronization transition.
Figure 1. Mean spike time tiling coefficient (STTC) as a function of coupling strength in a network of 36 coupled stochastic relaxation-type oscillators. Increasing coupling drives the system from weakly correlated spiking activity toward global synchronization, indicating a continuous synchronization transition.



