IntroductionSpatial navigation relies on integrating multimodal cues[1], yet both
in vivo and
in silico hippocampal research overwhelmingly focuses on vision [2,3,6]. While recent work showed mice entorhinal cortex contain both unimodal and multimodal cells[3], how different modalities are weighted and integrated remains poorly understood. We develop a modelling pipeline with an agent traversing a multimodal VR environment. A recurrent neural network was trained to perform a next-state prediction task[2]. We hypothesised that the network would develop place cell-like units that utilise both modalities, and that integrating modalities would result in more robust spatial representations, with each sense contributing additively to the cognitive map.
MethodsAn agent traversed a 2D arena with audiovisual cues in Unity3D[4]. Binaural audio broadcasted with head-related transfer functions[5] and visual frames were encoded via autoencoders into low-dimensional embeddings. We trained an RNN[2] to perform next-state prediction using its current sensory states and motion, under three conditions: audiovisual, visually-lesioned and auditorily-lesioned. Latent units were classified into place units using empirical metrics such as spatial information scores. These tunings were used to perform maximum-likelihood decoding of the agent’s position and orientation. The relative contributions of sensory inputs were effectively decomposed using a linearly weighted combination of their unimodal responses.
ResultsThe audiovisual model produced 127 spatially tuned place cells, significantly more than visually-driven (33 cells) and auditorily-driven (81 cells) ones. Furthermore, the audiovisual model yielded the lowest trajectory decoding error (0.151 m) compared to visual-only (0.879 m) and auditory-only (0.293 m) ones, with the highest spatial information content. Unimodal units that respond to a single modality were identified, as well as multimodal units that remap when both are present. Finally, by approximating multimodal ratemaps as linear combinations of unimodal maps, we found that most place cells integrate modalities additively, exhibiting intermediate visual weightings (μ=0.405) and relying more on auditory cues.
DiscussionWhile derived in silico, these results offer a framework for biological navigation. The model suggests the hippocampus may additively processes multisensory streams to reduce uncertainty rather than switching between senses. Notably, auditory cues proved dominant in our VR setup, likely because visual landmarks lose salience at a distance or vanish when facing walls. Consequently, multimodal units anchor to the most reliable available cues—in this case, sound. We further hypothesise these units will dynamically remap or reweigh sensory reliance if a primary modality degrades. Ultimately, this model provides a normative theory for multisensory integration, generating precise, testable predictions for planned in vivo ferret recordings.
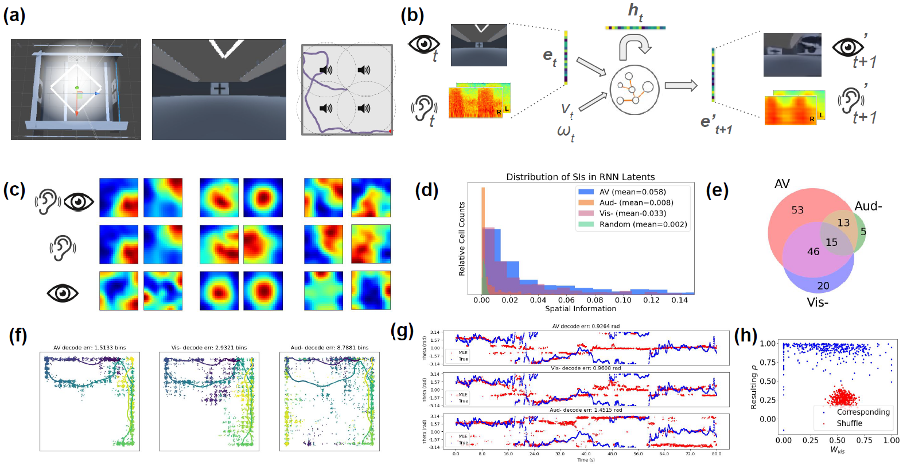 (a) Virtual environment with visual cues and sound sources; (b) Model architecture and pipeline; (c) Spatial ratemap examples in audiovisual, and lesioned (-) conditions; (d) Distribution of spatial information contents; (e) Number of place units identified; (f) ML decoding of position; (g) ML decoding of head direction; (h) Distribution of visual weights (x) and resulting correlations (y); comparReferences
(a) Virtual environment with visual cues and sound sources; (b) Model architecture and pipeline; (c) Spatial ratemap examples in audiovisual, and lesioned (-) conditions; (d) Distribution of spatial information contents; (e) Number of place units identified; (f) ML decoding of position; (g) ML decoding of head direction; (h) Distribution of visual weights (x) and resulting correlations (y); comparReferencesJeffery, K. J. (2007). Integration of the sensory inputs to place cells: what, where, why, and how?. Hippocampus
[1] Levenstein, D., ..., & Richards, B. (2024). Sequential predictive learning is a unifying theory for hippocampal representation and replay. bioRxiv
[2] Nguyen, D., ... , & Gu, Y. (2024). The medial entorhinal cortex encodes multisensory spatial information. Cell reports
[3] George, T. M., ..., & Barry, C. (2024). RatInABox, a toolkit for modelling locomotion and neuronal activity in continuous environments. Elife
[4] Cuevas-Rodríguez, ... & Reyes-Lecuona, A. (2019). 3D Tune-In Toolkit: An open-source library for real-time binaural spatialisation. PloS one
[5] Banino, A., ... & Kumaran, D. (2018). Vector-based navigation using grid-like representations in artificial agents. Nature
AcknowledgementWe thank Barry Lab, Bizley La, the Department of UCL Cell and Developmental Biology, the Ear Institute for this work.
This work was supported by the UKRI Biotechnology and Biological Sciences Research Council [grant
number BB/T008709/1].
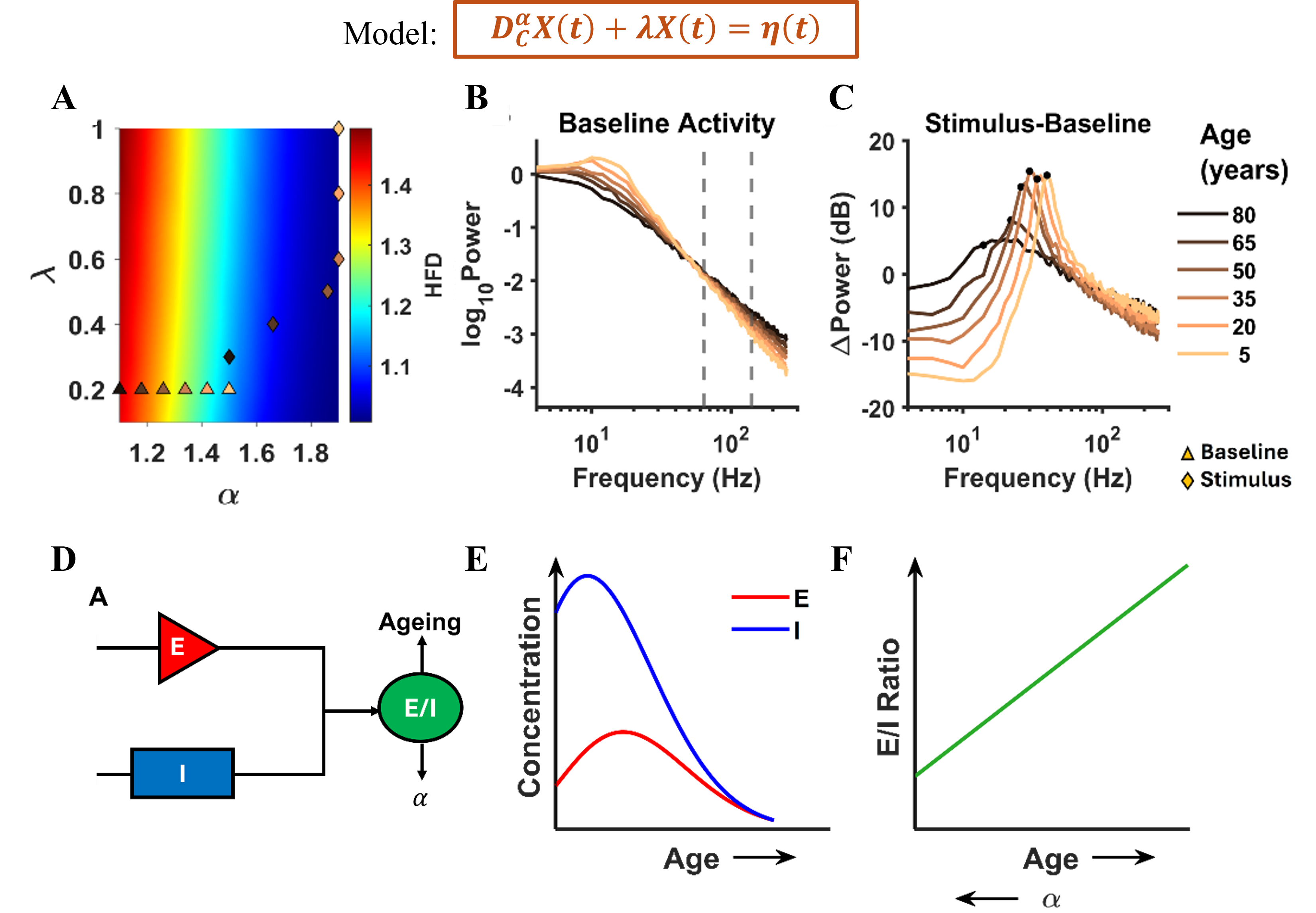 Figure 1. Age-related inferences: Top: Model equation. (A) HFD variation with α and λ. (B), (C) The PSDs and ΔPower (in dB) corresponding to triangles and diamonds respectively. (D) Illustration of E/I dependence on ageing and α. (E) The concentration of excitatory and inhibitory neurotransmitters varying non-monotonically with age. (F) Corresponding monotonic E/I ratio with age and α.
Figure 1. Age-related inferences: Top: Model equation. (A) HFD variation with α and λ. (B), (C) The PSDs and ΔPower (in dB) corresponding to triangles and diamonds respectively. (D) Illustration of E/I dependence on ageing and α. (E) The concentration of excitatory and inhibitory neurotransmitters varying non-monotonically with age. (F) Corresponding monotonic E/I ratio with age and α.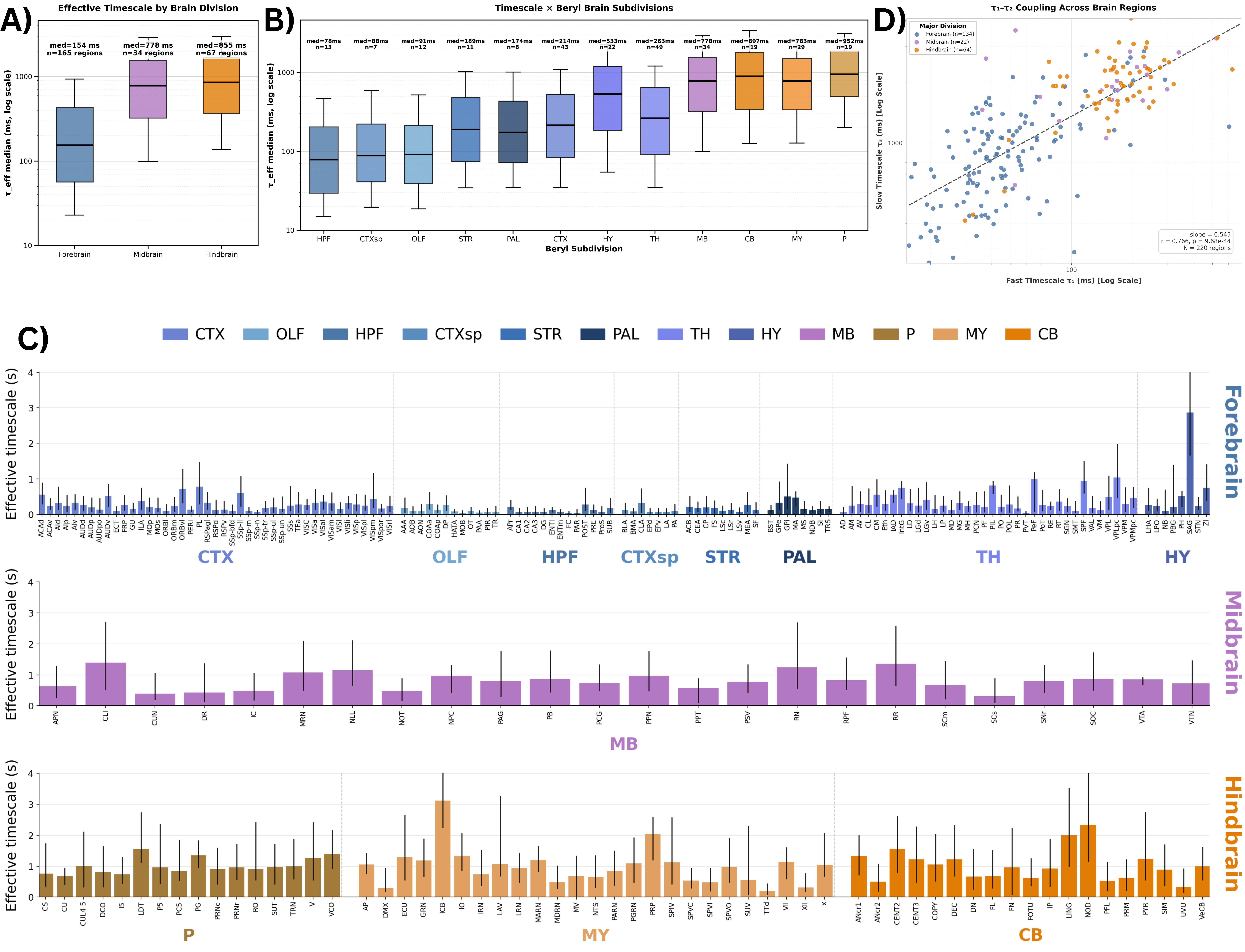 Brain-wide map of intrinsic neural timescales. (A) τ_eff by major brain division. (B) τ_eff across 12 brain subdivisions. (C) Median τ_eff per region; error bars show 10th–90th percentiles. Regions grouped by division, ordered alphabetically; colors denote subdivision. (D) Fast (τ₁) vs slow (τ₂) timescales across regions.
Brain-wide map of intrinsic neural timescales. (A) τ_eff by major brain division. (B) τ_eff across 12 brain subdivisions. (C) Median τ_eff per region; error bars show 10th–90th percentiles. Regions grouped by division, ordered alphabetically; colors denote subdivision. (D) Fast (τ₁) vs slow (τ₂) timescales across regions.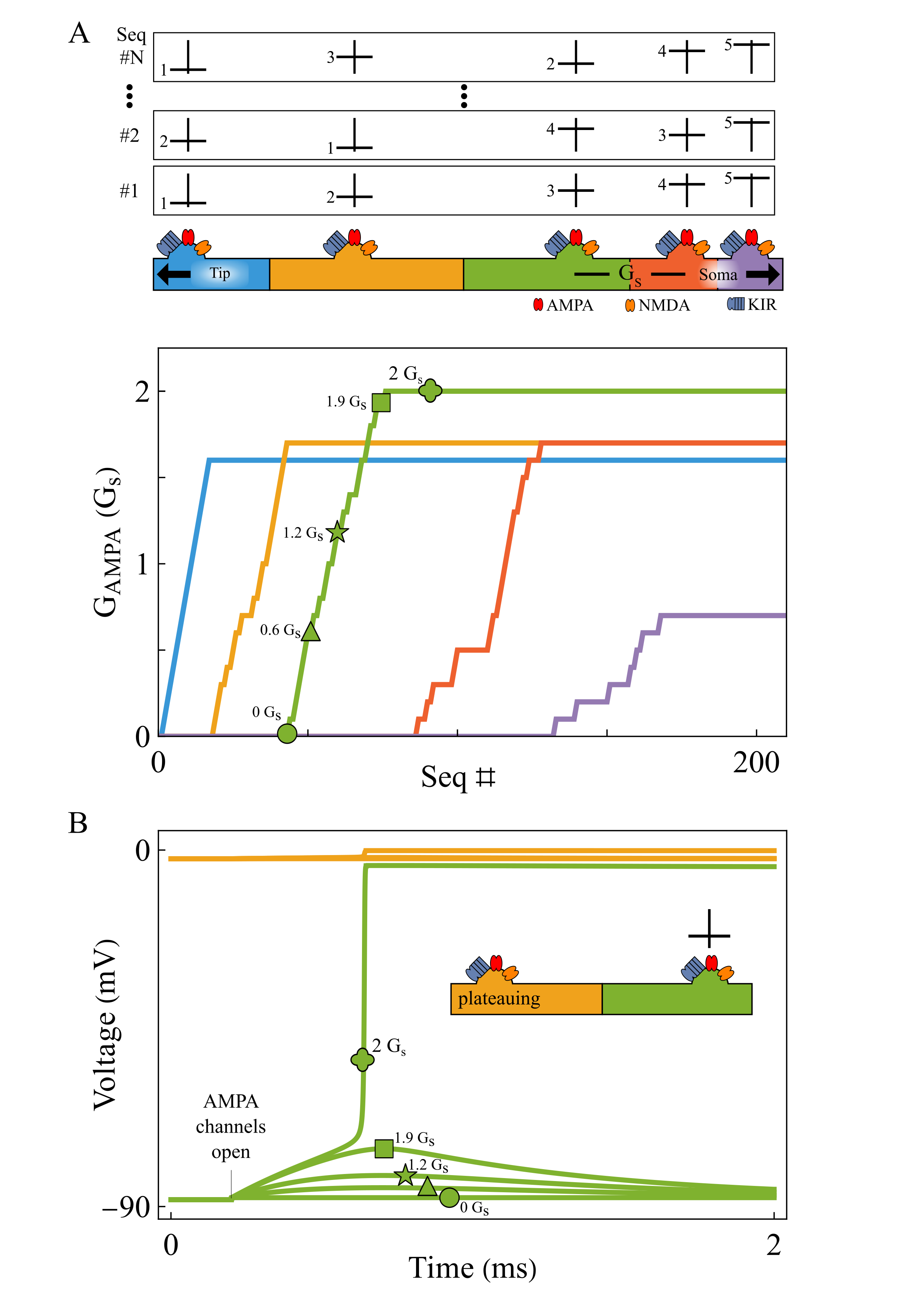 Local AMPA plasticity tunes dendritic sequence selectivity. Repeated tip-to-soma and shuffled inputs progressively strengthen AMPA conductance from tip to soma (A). When a segment (green) receives input while its tip-side neighbor is plateauing (yellow), weak AMPA initially fails to trigger a plateau. As AMPA increases, the segment abruptly transitions to plateau at a bifurcation (B).References
Local AMPA plasticity tunes dendritic sequence selectivity. Repeated tip-to-soma and shuffled inputs progressively strengthen AMPA conductance from tip to soma (A). When a segment (green) receives input while its tip-side neighbor is plateauing (yellow), weak AMPA initially fails to trigger a plateau. As AMPA increases, the segment abruptly transitions to plateau at a bifurcation (B).References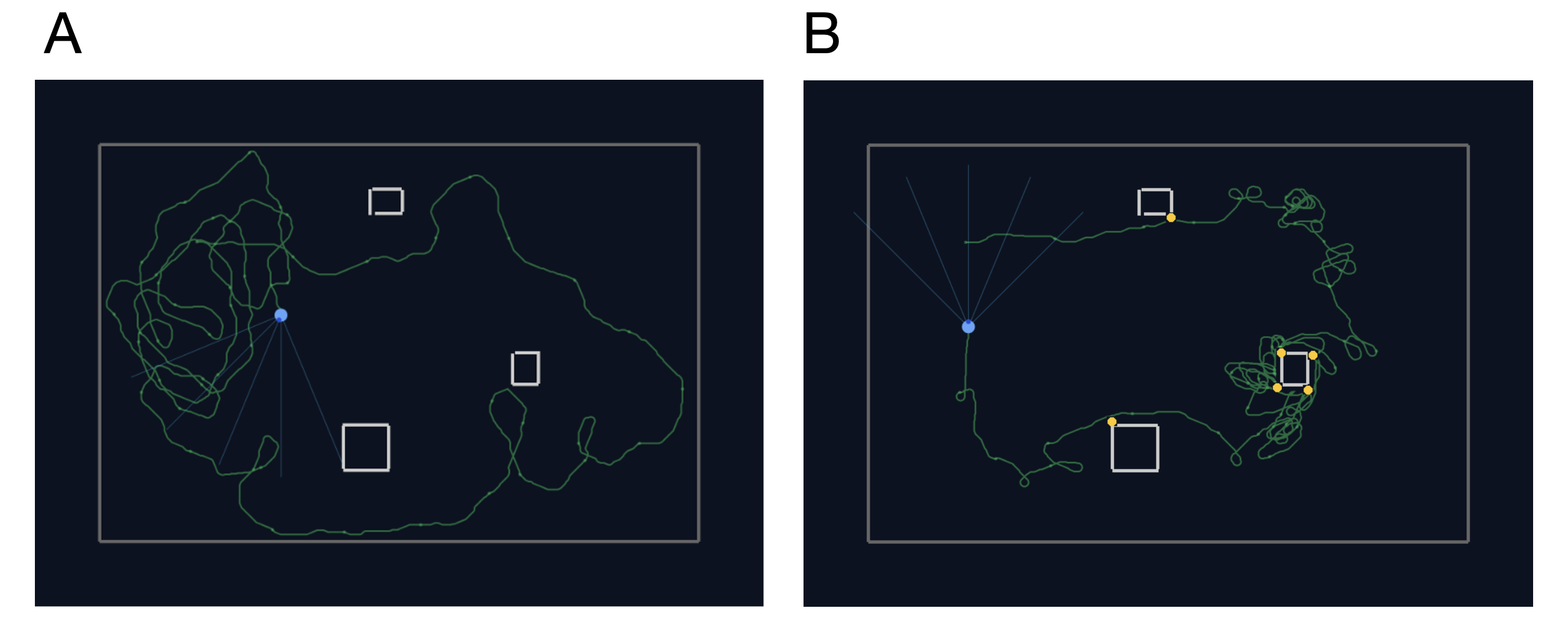 Example trajectories (green) of an agent (light blue circle with blue rays indicating visual sensors) in an arena with three high-contrast obstacles. A: The agent forages where no obstacles are present. It also frequents the part of the arena with obstacles, but never collides with them. B: Trajectory for an agent with less training times, leading to frequent collisions (yellow dots).References
Example trajectories (green) of an agent (light blue circle with blue rays indicating visual sensors) in an arena with three high-contrast obstacles. A: The agent forages where no obstacles are present. It also frequents the part of the arena with obstacles, but never collides with them. B: Trajectory for an agent with less training times, leading to frequent collisions (yellow dots).References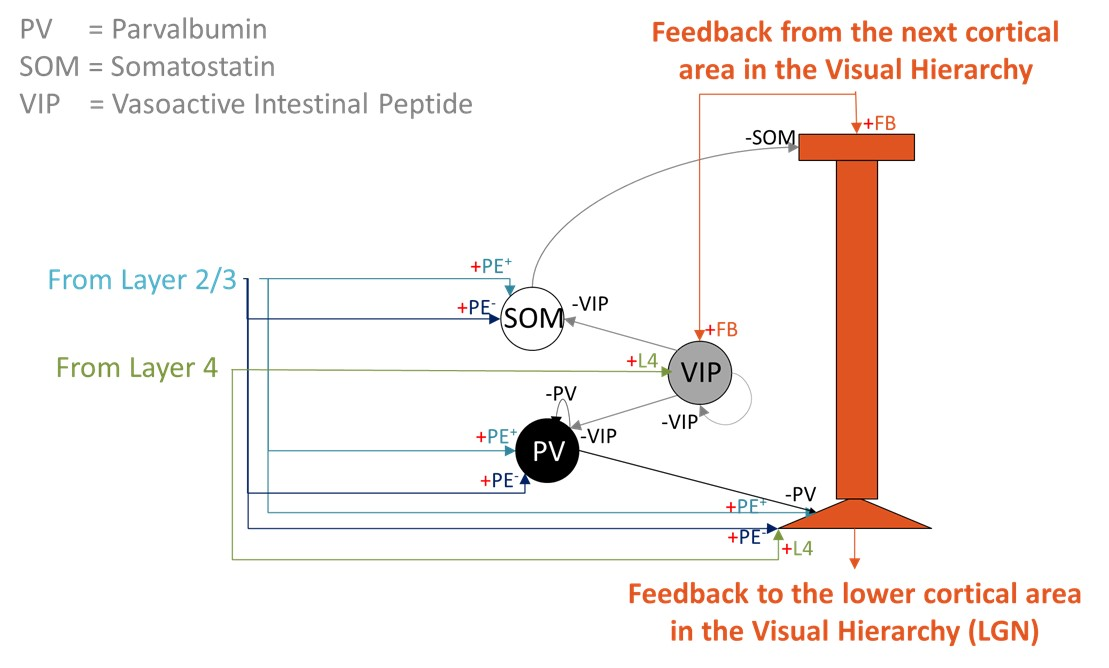 Layer 5 predictive-coding microcircuit. A two-compartment Layer 5 pyramidal neuron integrates somatic input from Layer 2/3 prediction-error populations (PE+ and PE-) and Layer 4 feature neurons with apical feedback from higher cortical areas. A local VIP-PV-SOM motif regulates apical Ca2+ spikes and burst output.
Layer 5 predictive-coding microcircuit. A two-compartment Layer 5 pyramidal neuron integrates somatic input from Layer 2/3 prediction-error populations (PE+ and PE-) and Layer 4 feature neurons with apical feedback from higher cortical areas. A local VIP-PV-SOM motif regulates apical Ca2+ spikes and burst output.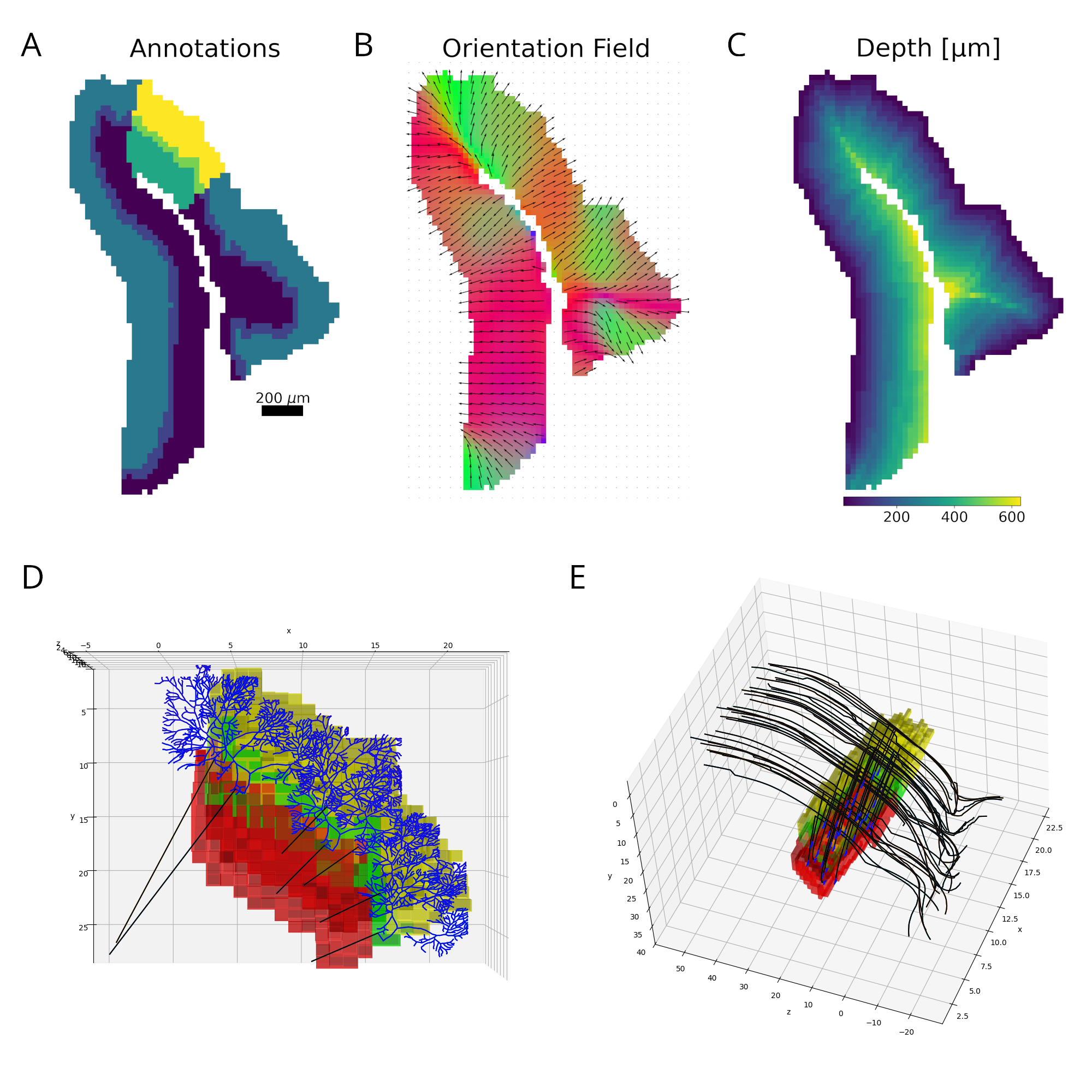 Figure 1. Reconstruction pipeline. A. Declive layers shown in colors with the selected column highlighted. B. Orientation field showing the local axons’ main axis. Colors represent the vectors’ norm. C. Distance to the outside border, following the orientation field. D. E. Purkinje and granule cells´ morphology scaled and bent according to the declive shape.
Figure 1. Reconstruction pipeline. A. Declive layers shown in colors with the selected column highlighted. B. Orientation field showing the local axons’ main axis. Colors represent the vectors’ norm. C. Distance to the outside border, following the orientation field. D. E. Purkinje and granule cells´ morphology scaled and bent according to the declive shape.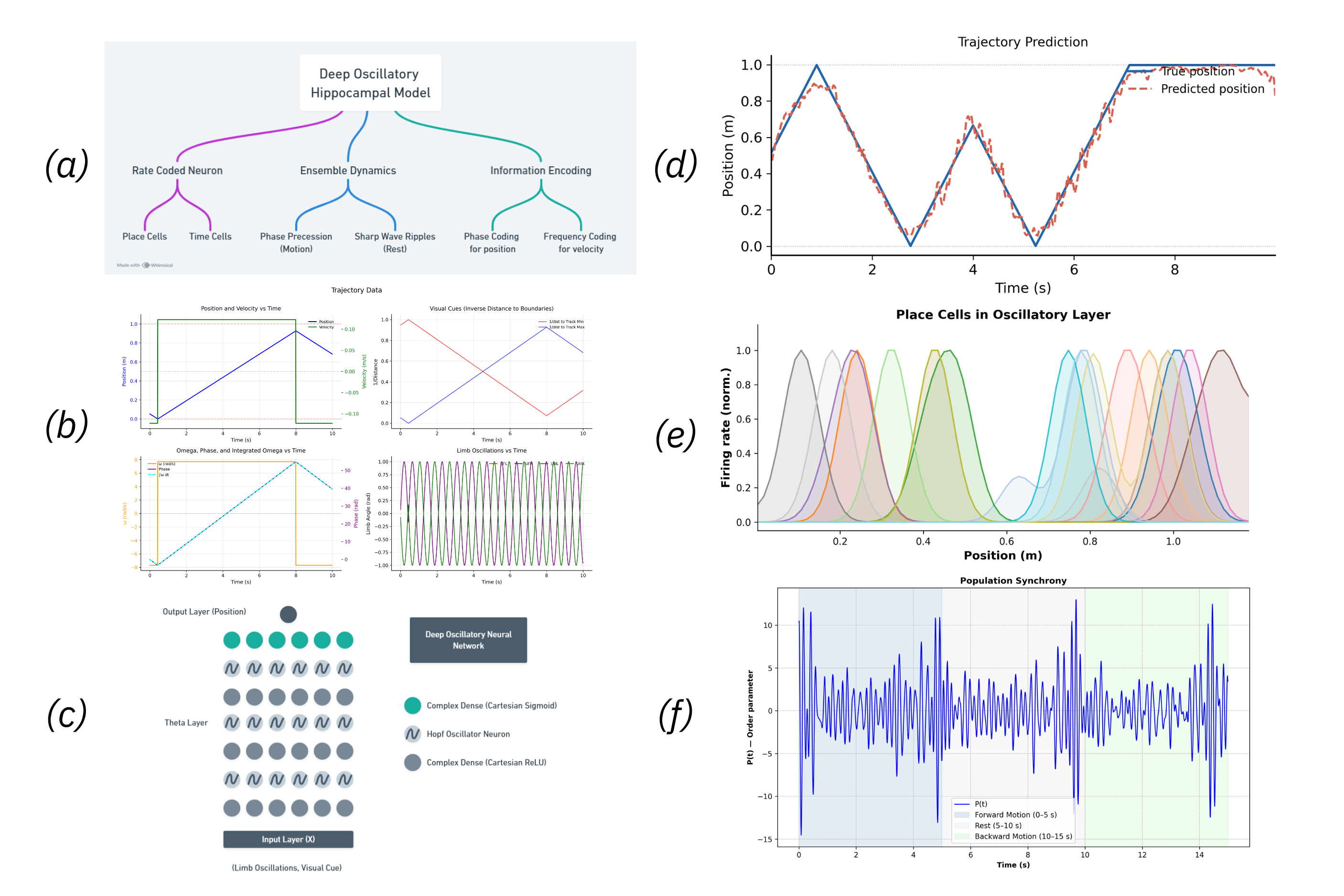 Figure 1. (a) Model Flowchart, (b) Input Data, (c) Oscillatory Neural Network Diagram, (d) Trajectory Prediction, (e) Place Cells - different colors correspond to different neurons, (f) Sharp Wave Ripples
Figure 1. (a) Model Flowchart, (b) Input Data, (c) Oscillatory Neural Network Diagram, (d) Trajectory Prediction, (e) Place Cells - different colors correspond to different neurons, (f) Sharp Wave Ripples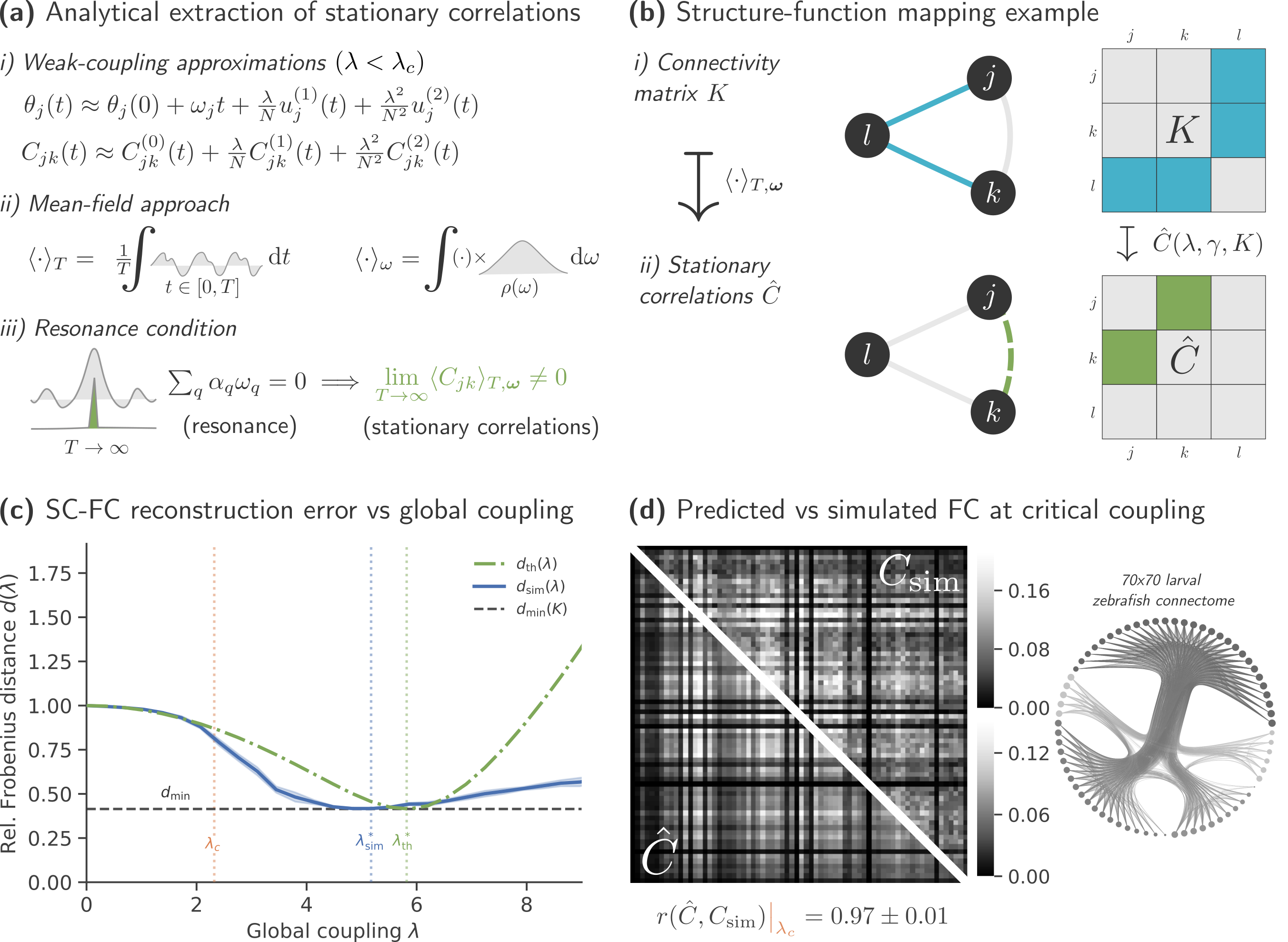 Figure 1. Predicting coactivity from anatomy in neural oscillators. (a) Derivation of predicted functional connectivity: phase trajectories are expanded, correlations averaged, and stationary terms selected. (b) Example for N=2 oscillators. (c) SC–FC reconstruction error follows theory up to synchronization (λc = 2.32). (d) Predicted and simulated FC remain highly similar (cosine similarity ≥ 0.97).
Figure 1. Predicting coactivity from anatomy in neural oscillators. (a) Derivation of predicted functional connectivity: phase trajectories are expanded, correlations averaged, and stationary terms selected. (b) Example for N=2 oscillators. (c) SC–FC reconstruction error follows theory up to synchronization (λc = 2.32). (d) Predicted and simulated FC remain highly similar (cosine similarity ≥ 0.97).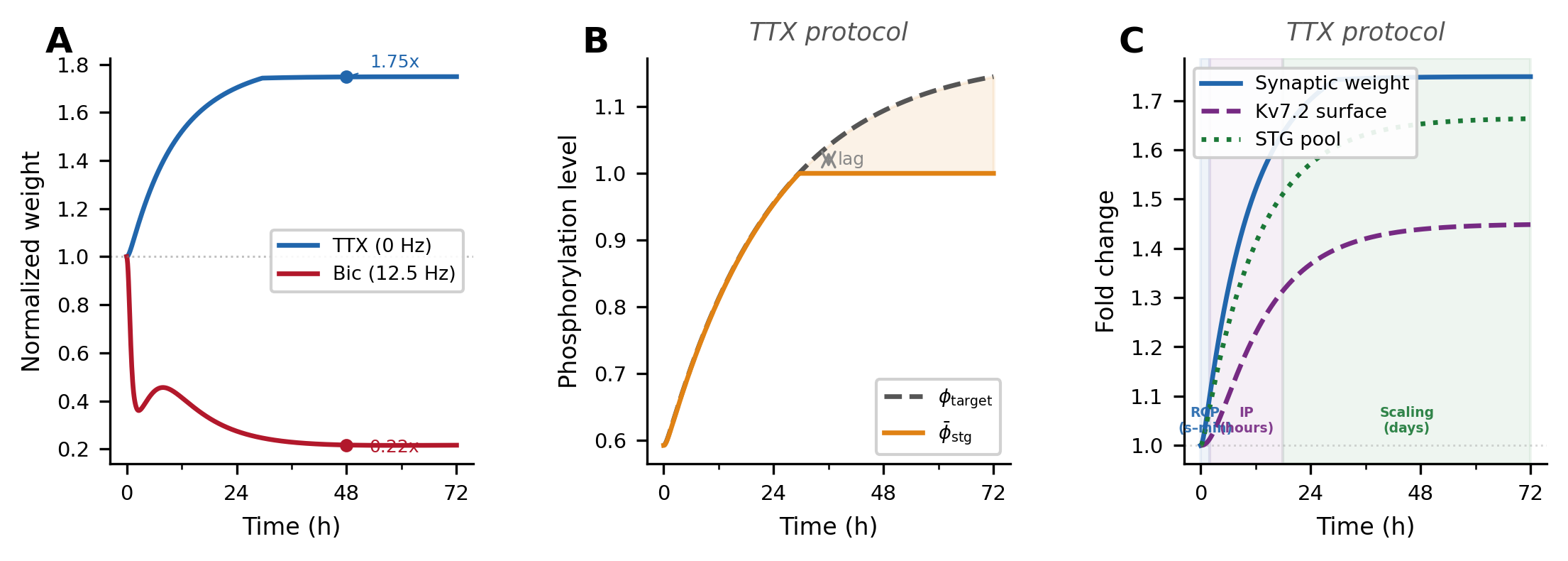 Figure 1. Simulated homeostatic response to inactivity (TTX). (A) Multiplicative synaptic scaling: upscaling (1.75×) and downscaling (0.22×) at 48 h. (B) Stargazin phosphorylation (φ̄_stg) lags the homeostatic target (φ_target) due to enzymatic inertia. (C) Three compensatory tiers emerge sequentially: AMPAR redistribution (seconds–min), Kv7.2 adjustment (hours), and synaptic scaling (days).
Figure 1. Simulated homeostatic response to inactivity (TTX). (A) Multiplicative synaptic scaling: upscaling (1.75×) and downscaling (0.22×) at 48 h. (B) Stargazin phosphorylation (φ̄_stg) lags the homeostatic target (φ_target) due to enzymatic inertia. (C) Three compensatory tiers emerge sequentially: AMPAR redistribution (seconds–min), Kv7.2 adjustment (hours), and synaptic scaling (days).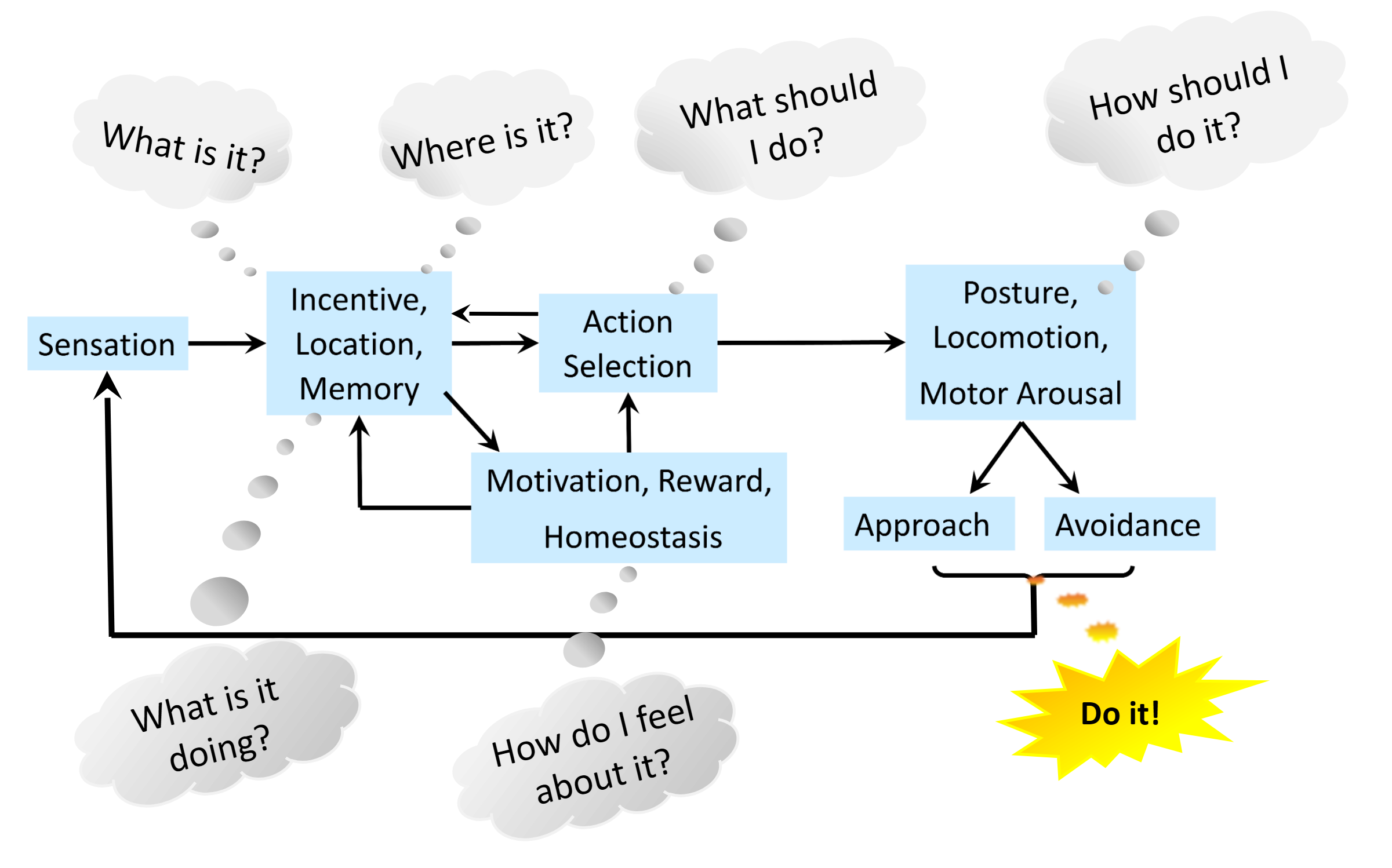 Figure 1. Both simple and complex animals handle flow of information from sensation to behavior with a common modular nervous system organization. Stimuli characteristics, incentives and locations are integrated with memory, motivation and affect for decisive action selection, with 5 critical computations from “What is it?” to “How should I do it?”, and cognitive mapping adding “What is it doing?”.
Figure 1. Both simple and complex animals handle flow of information from sensation to behavior with a common modular nervous system organization. Stimuli characteristics, incentives and locations are integrated with memory, motivation and affect for decisive action selection, with 5 critical computations from “What is it?” to “How should I do it?”, and cognitive mapping adding “What is it doing?”.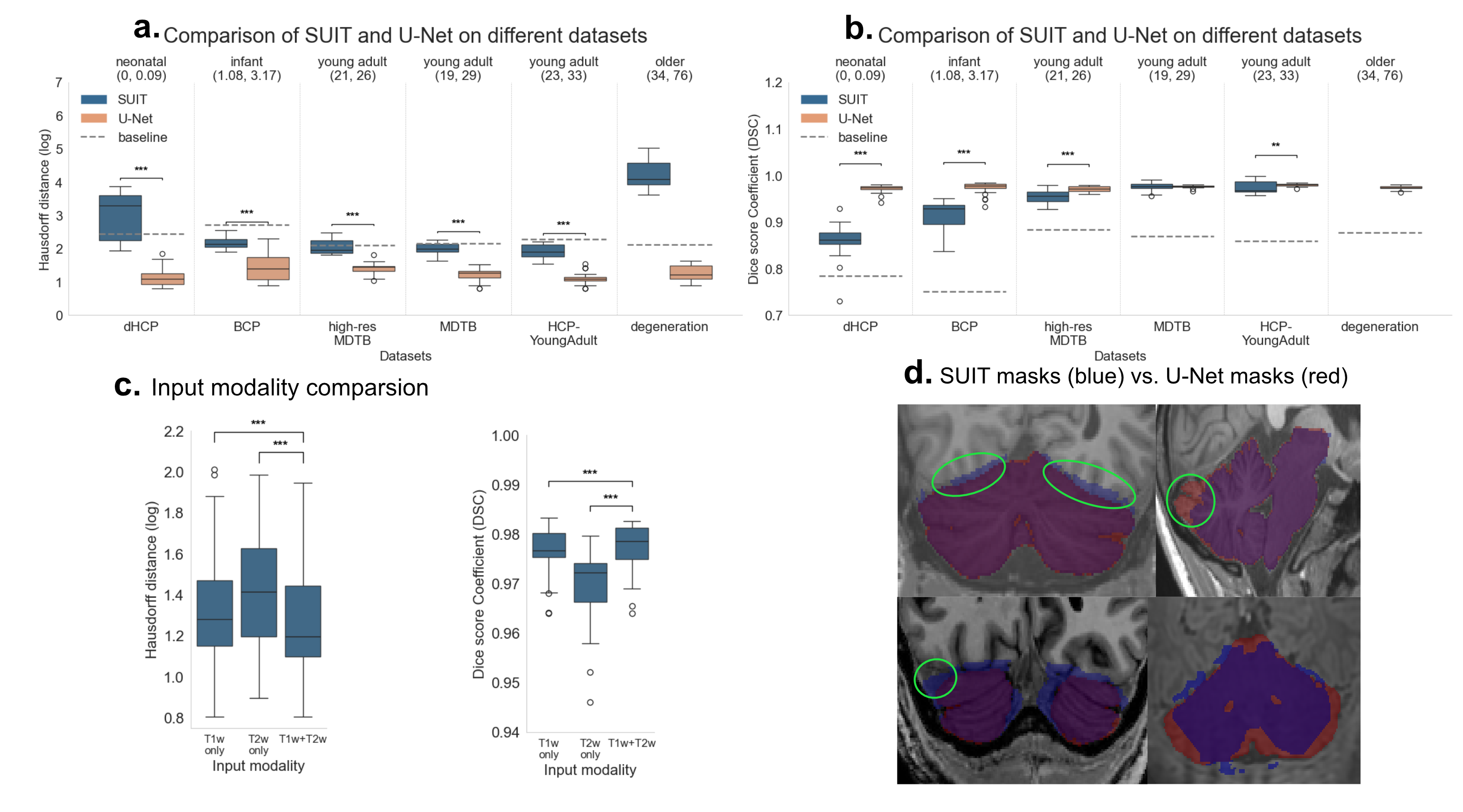 Figure 1. Isolation analysis. Comparison of U-Net VS SUIT for (a) Hausdorff Distance (HD) and (b) Dice Score Coefficient (DSC). c shows performance for different input modalities in the full datasets. Horizontal lines between bars with asterisks denote significant differences (paired t-tests). Baseline: Average mask prediction. d shows resulting mask in problematic subjects.
Figure 1. Isolation analysis. Comparison of U-Net VS SUIT for (a) Hausdorff Distance (HD) and (b) Dice Score Coefficient (DSC). c shows performance for different input modalities in the full datasets. Horizontal lines between bars with asterisks denote significant differences (paired t-tests). Baseline: Average mask prediction. d shows resulting mask in problematic subjects.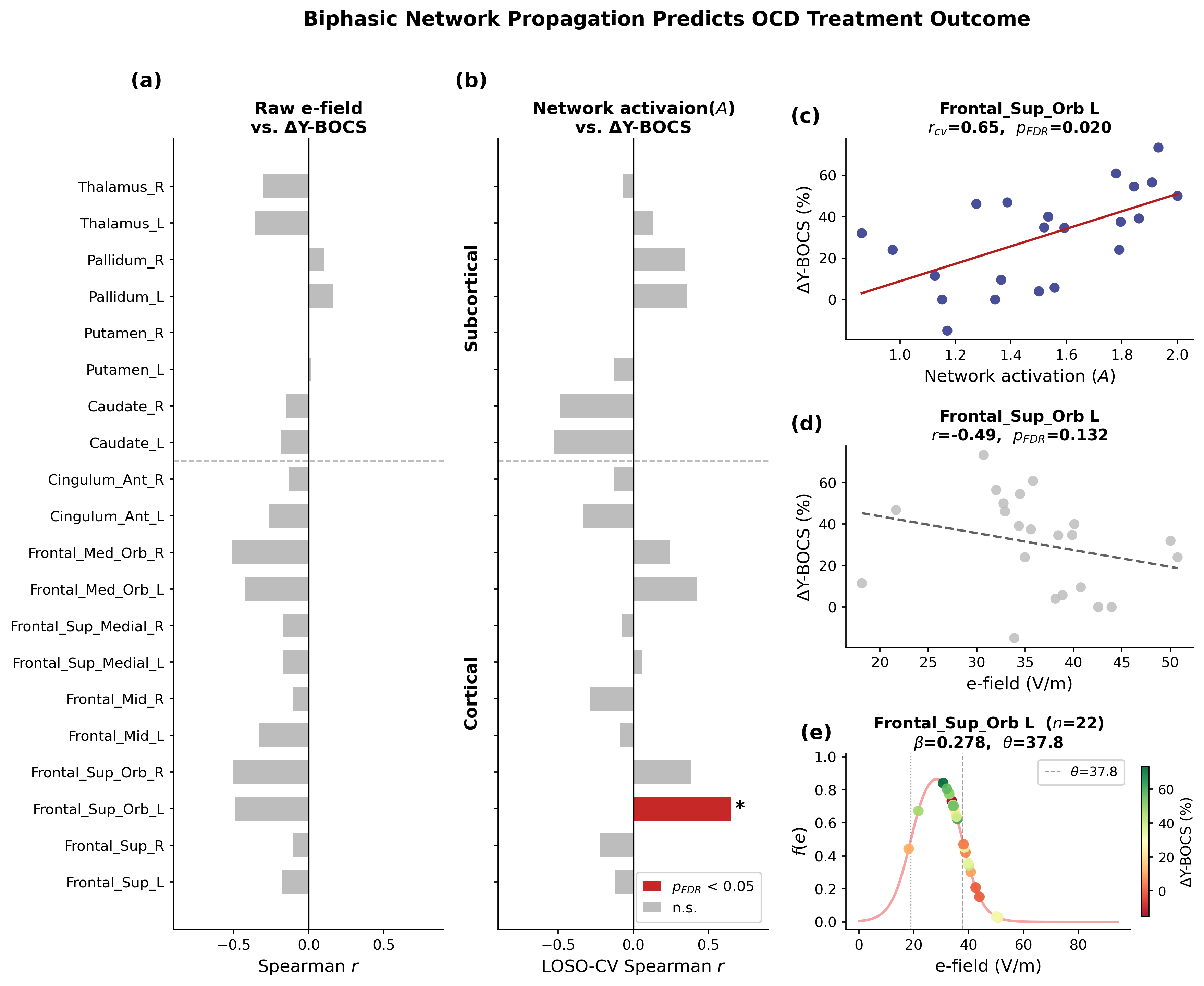 Figure 1. Correlations between E-field/network activation and clinical outcome in the CSTC ROIs. (a) Raw E-field showed no significant correlations. (b) The biphasic neuronal response model identified the left superior orbital frontal cortex as significantly associated with percentage Y-BOCS improvement. (c-f) Network activation/raw E-field correlation maps, and fitted biphasic neuronal response fReferences
Figure 1. Correlations between E-field/network activation and clinical outcome in the CSTC ROIs. (a) Raw E-field showed no significant correlations. (b) The biphasic neuronal response model identified the left superior orbital frontal cortex as significantly associated with percentage Y-BOCS improvement. (c-f) Network activation/raw E-field correlation maps, and fitted biphasic neuronal response fReferences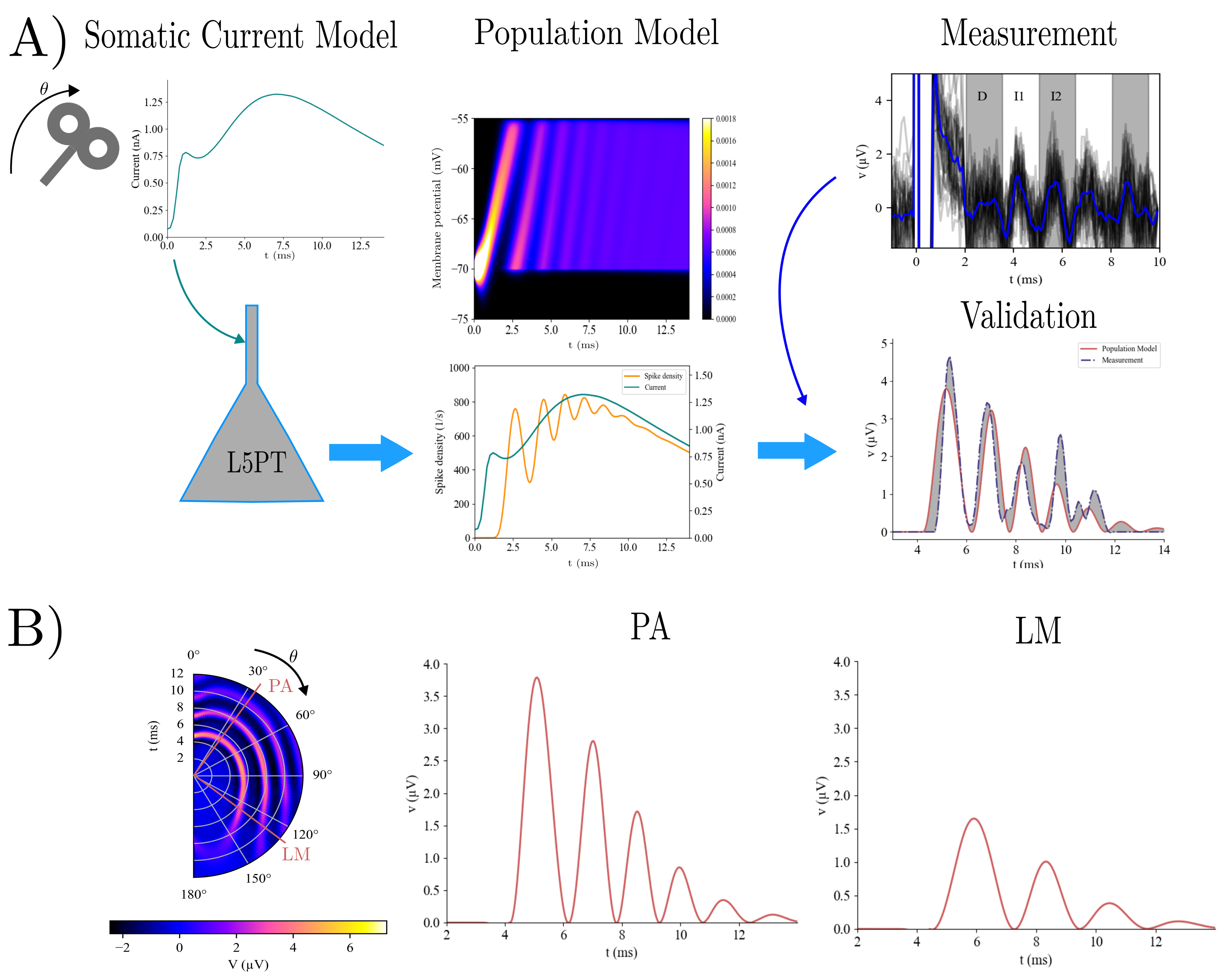 Figure 1. A) The somatic current model [3] generates coil orientation-sensitive somatic currents (first column). They are then applied to the L5PT model which computes membrane potential distribution and spike density for it (second column). This is then transformed to a potential and compared to measured I-waves. B) Orientation dependency of I-waves (first column) and potentials for putative parietal-anter
Figure 1. A) The somatic current model [3] generates coil orientation-sensitive somatic currents (first column). They are then applied to the L5PT model which computes membrane potential distribution and spike density for it (second column). This is then transformed to a potential and compared to measured I-waves. B) Orientation dependency of I-waves (first column) and potentials for putative parietal-anter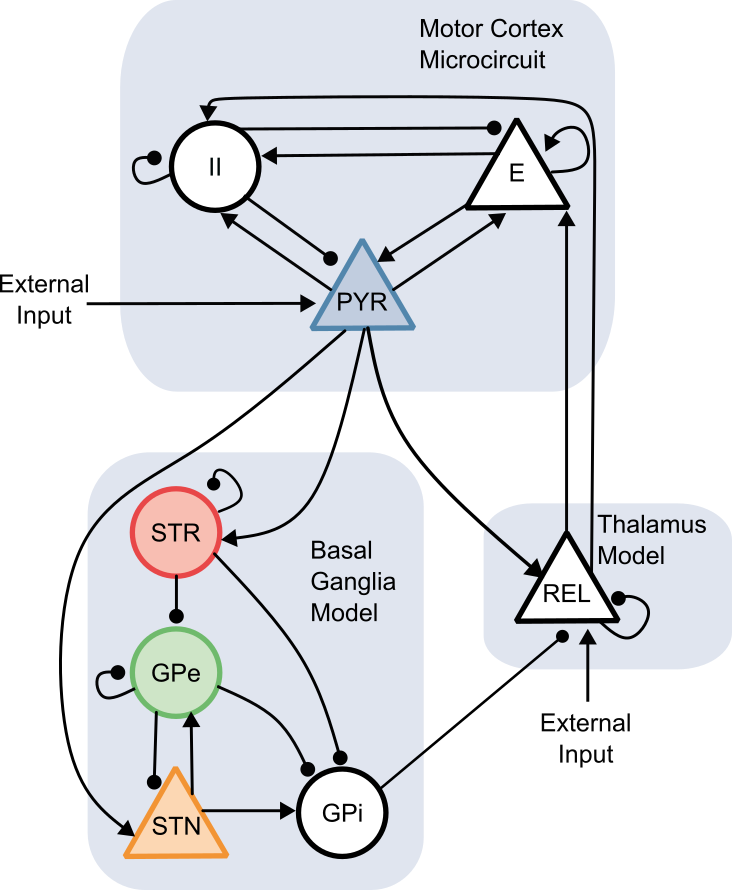 Figure 1. The BGTC circuit connectivity implemented in the neural mass model includes cortical excitatory (E), inhibitory interneuron (II), and deep pyramidal (DP) populations, as well as the striatum, globus pallidus externus (GPe), globus pallidus internus (GPi), subthalamic nucleus (STN), and thalamic relay nuclei (REL). Shaded regions indicate nodes with spectral data used for model fitting.
Figure 1. The BGTC circuit connectivity implemented in the neural mass model includes cortical excitatory (E), inhibitory interneuron (II), and deep pyramidal (DP) populations, as well as the striatum, globus pallidus externus (GPe), globus pallidus internus (GPi), subthalamic nucleus (STN), and thalamic relay nuclei (REL). Shaded regions indicate nodes with spectral data used for model fitting.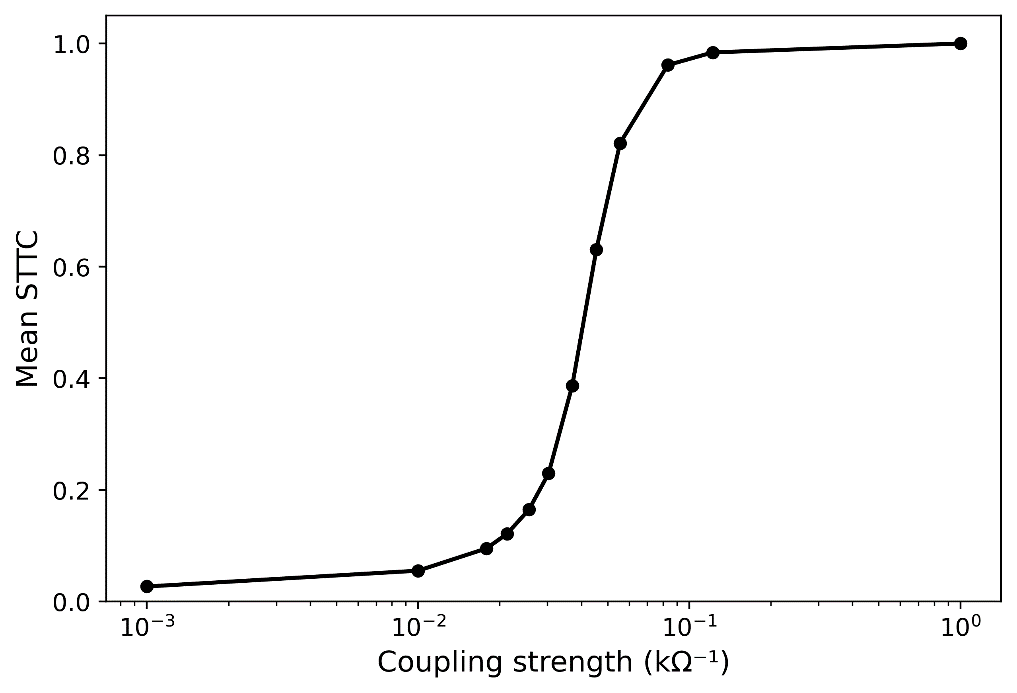 Figure 1. Mean spike time tiling coefficient (STTC) as a function of coupling strength in a network of 36 coupled stochastic relaxation-type oscillators. Increasing coupling drives the system from weakly correlated spiking activity toward global synchronization, indicating a continuous synchronization transition.
Figure 1. Mean spike time tiling coefficient (STTC) as a function of coupling strength in a network of 36 coupled stochastic relaxation-type oscillators. Increasing coupling drives the system from weakly correlated spiking activity toward global synchronization, indicating a continuous synchronization transition.



