Hannah van Susteren1,*, Guillaume Girier
2,*, Michel J.A.M. van Putten
3,4 , Jaroslav Hlinka
2, Helmut Schmidt
2, Hil G.E. Meijer
1
1 Department of Applied Mathematics, University of Twente, Enschede, the Netherlands
2 Institute of Computer Science, Czech Academy of Sciences, Prague, Czech Republic
3 Department of Neurology and Clinical Neurophysiology, University of Twente, Enschede, the Netherlands
4 Medisch Spectrum Twente, Enschede, the Netherlands
* These authors contributed equally to this work.
Email:
[email protected]
IntroductionEpilepsy is among the most prevalent neurological disorders. The astrocytic excitatory amino acid transporter (EAAT2) plays a key role in regulating excitability, by controlling extracellular glutamate levels and glutamate receptor activation [1,2]. Reduced EAAT2 expression has been reported in several epilepsy patients [1,3]. However, the impact of neuron-astrocyte interactions on hyperexcitability on single cell level is underexplored. We developed a biophysical model of a presynaptic neuron and astrocyte to explore the relation between astrocytic EAAT2-mediated glutamate clearance, presynaptic glutamate receptors and bursting activity.MethodsWe build on our previous work [4,5], where we consider a presynaptic neuron and an astrocyte in a finite extracellular space (ECS). This model describes sodium, potassium, chloride dynamics as well as calcium-dependent exocytosis and glutamate-glutamine (GG) recycling. For this study, we add a potassium bath with diffusion to the ECS to induce neuronal bursting (Fig. 1A). Additionally, we implement the presynaptic glutamate receptors AMPA and NMDA, which are important in regulating hyperexcitability. Lastly, we study the impact of the antiseizure drugs perampanel and memantine by simulating the effect of these AMPA and NMDA receptor antagonists.ResultsWe induce neuronal bursting by increasing extracellular potassium in the bath. We first examine how AMPA and NMDA permeabilities affect burst frequency (Fig. 1C), where frequency refers to spike frequency during the last burst or during tonic firing, to fit the NMDA/AMPA ratio to experimental data [6]. Higher permeabilities increase neuronal firing and intracellular calcium, triggering a feedback loop that enhances neuronal glutamate release. Reducing EAAT permeability raises burst frequency and induces tonic firing (Fig. 1B). Finally, AMPA and NMDA antagonists, perampanel and memantine [7], reduce firing despite elevated extracellular glutamate, with perampanel showing a more significant reduction in firing frequency (Fig. 1D). DiscussionOur results show that reduced EAAT expression, as observed in several epilepsy patients, results in increased extracellular glutamate and overstimulation of excitatory glutamate receptors. Furthermore, we show that the AMPA and NMDA receptor permeabilities affect burst frequency. Receptor antagonists such as perampanel and memantine are able to reduce firing. In conclusion, our detailed neuron–astrocyte model provides insight into the effects of reduced EAAT expression and receptor antagonists on hyperexcitability.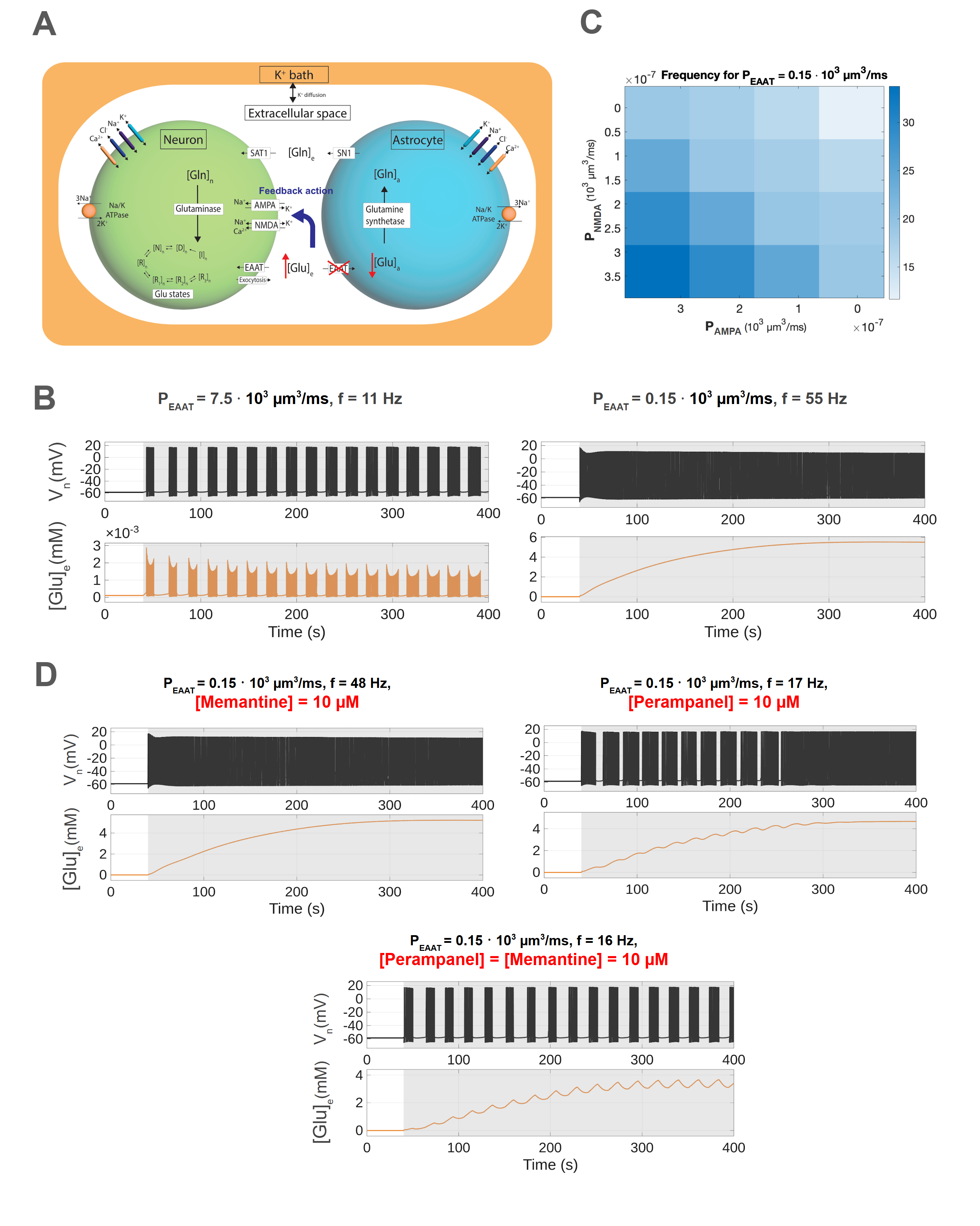 Figure 1. A: Three-compartment model illustrating the GG-cycle during EAAT2 knockout. B: The membrane potential, spike frequency f and ECS glutamate at different EAAT2 permeabilities. C: Spike frequency within bursts as a function of NMDA and AMPA receptor permeability. D: Neuronal activity at fixed EAAT2 permeability (PEAAT=0.15 * 103 µm3/ms) under antagonist conditions.References
Figure 1. A: Three-compartment model illustrating the GG-cycle during EAAT2 knockout. B: The membrane potential, spike frequency f and ECS glutamate at different EAAT2 permeabilities. C: Spike frequency within bursts as a function of NMDA and AMPA receptor permeability. D: Neuronal activity at fixed EAAT2 permeability (PEAAT=0.15 * 103 µm3/ms) under antagonist conditions.References[1] Green, J. L., dos Santos, W. F., & Fontana, A. C. K. (2021). Biochemical Pharmacology, 10.1016/j.bcp.2021.114786
[2] Scimemi, A., Tian, H., & Diamond, J. S. (2009). The Journal of Neuroscience, 10.1523/JNEUROSCI.4845-09.2009
[3] Barker-Haliski, M., & White, H. (2015). Cold Spring Harbor perspectives in medicine, 10.1101/cshperspect.a022863
[4] van Susteren, H., Rose, C. R., van Putten, M. J., & Meijer, H. G. (2025). bioRxiv, 10.1101/2025.11.10.687543
[5] Kalia, M., et al. (2021). PLOS Computational Biology, 10.1371/journal.pcbi.1009019
[6] Watt, A. J., Sjöström, P. J., Häusser, M., Nelson, S. B., & Turrigiano, G. G. (2004). Nature neuroscience, 10.1038/nn1220
[7] Chen, T.-S., Huang, T.-H., Lai, M.-C., & Huang, C.-W. (2023). Biomedicines, 10.3390/biomedicines11030783
AcknowledgmentsHVS, HGEM, MJAMVP funded from the DFG, FOR2795 ‘Synapses under stress’ to CRR (Prof. Dr. Christine R. Rose) (Ro2327/13-2 and 14-2).
GG, HS, and JH were supported by the ERDF-Project Brain dynamics, No. CZ.02.01.01/00/22\_008/0004643, a Lumina-Quaeruntur fellowship (LQ100302301), and the long-term strategic development financing of the Institute of Computer Science (RVO:67985807).
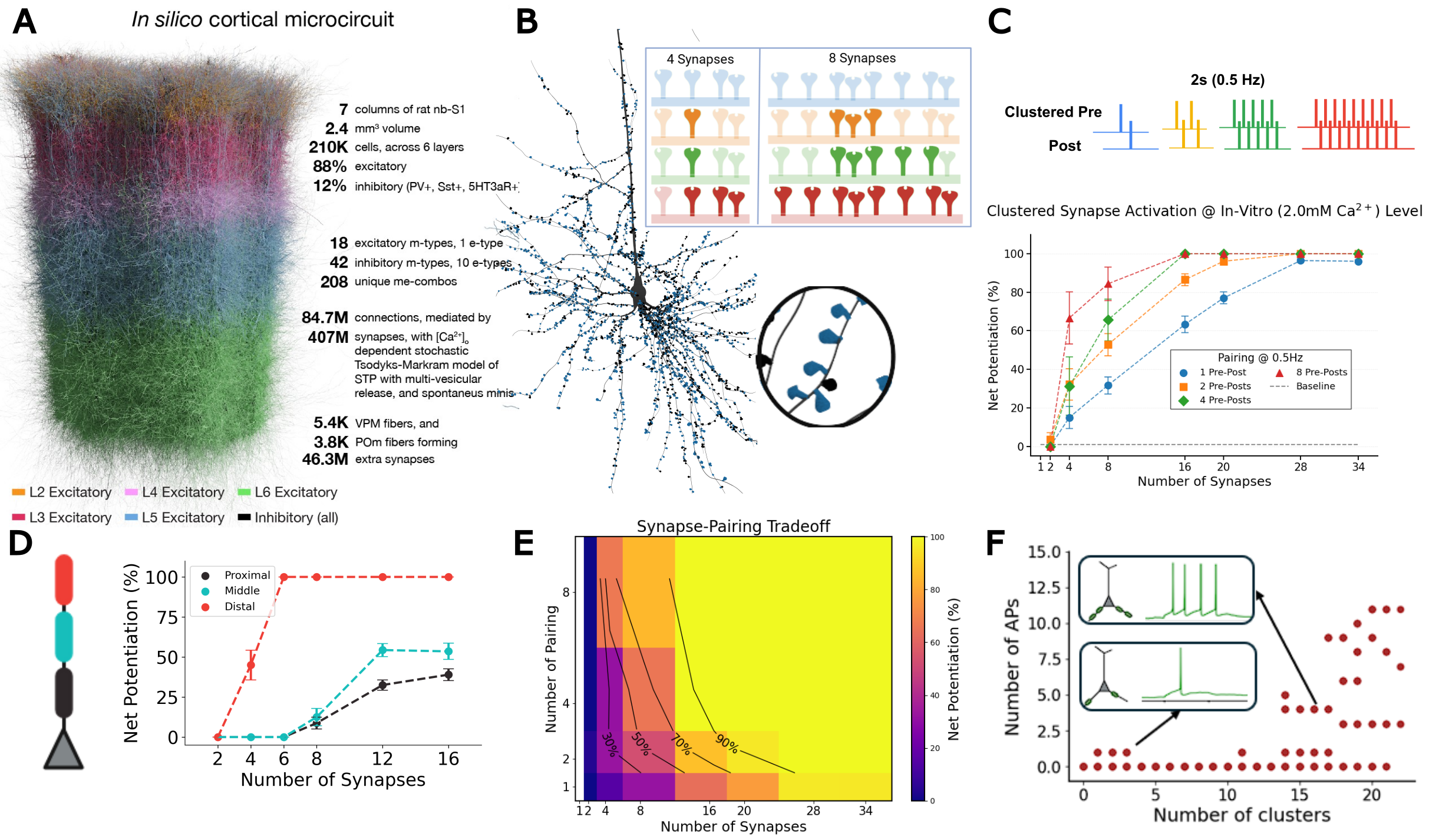 Figure 1. A, In silico cortical microcircuit. B, L5-TTPC with magnified cluster showing plasticity for 4 vs 8 co-active synapses. C, Clustered pre-post pairing (0.5 Hz); net potentiation vs synapse number. D, Spatial learning gradient E, Synapse-pairing tradeoff heatmap. F, Basal cluster coactivation triggers somatic burst.
Figure 1. A, In silico cortical microcircuit. B, L5-TTPC with magnified cluster showing plasticity for 4 vs 8 co-active synapses. C, Clustered pre-post pairing (0.5 Hz); net potentiation vs synapse number. D, Spatial learning gradient E, Synapse-pairing tradeoff heatmap. F, Basal cluster coactivation triggers somatic burst.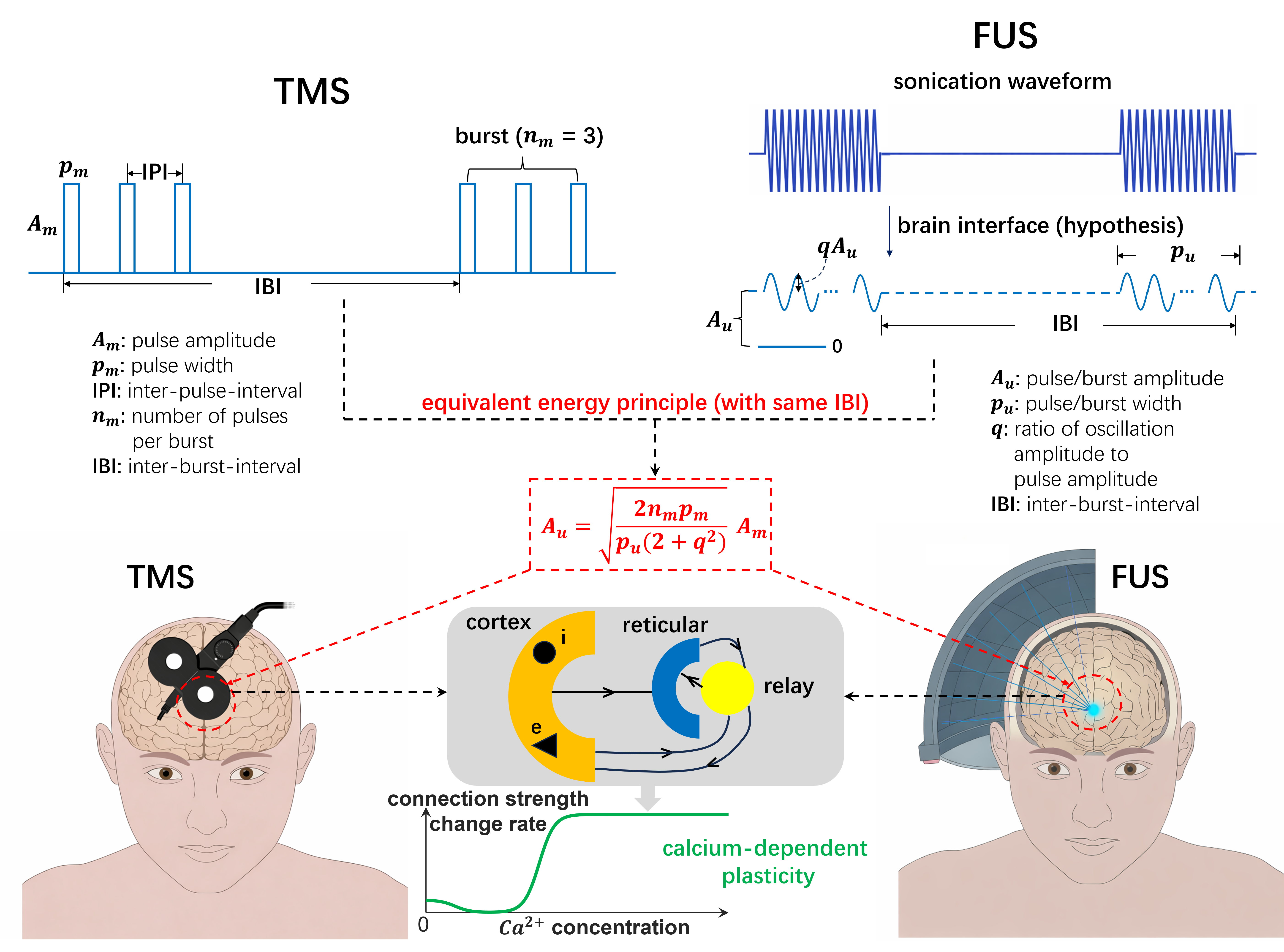 Figure 1. Equivalent energy principle for aligning rTMS and rTFUS plasticity models. rTMS delivers discrete pulse bursts; rTFUS delivers continuous bursts filtered by the skull interface to sub-1kHz sinusoids. The principle constrains parameters so both modalities deliver equal energy at the same carrier frequency, enabling unified mean-field modelling of calcium-dependent plasticity.
Figure 1. Equivalent energy principle for aligning rTMS and rTFUS plasticity models. rTMS delivers discrete pulse bursts; rTFUS delivers continuous bursts filtered by the skull interface to sub-1kHz sinusoids. The principle constrains parameters so both modalities deliver equal energy at the same carrier frequency, enabling unified mean-field modelling of calcium-dependent plasticity.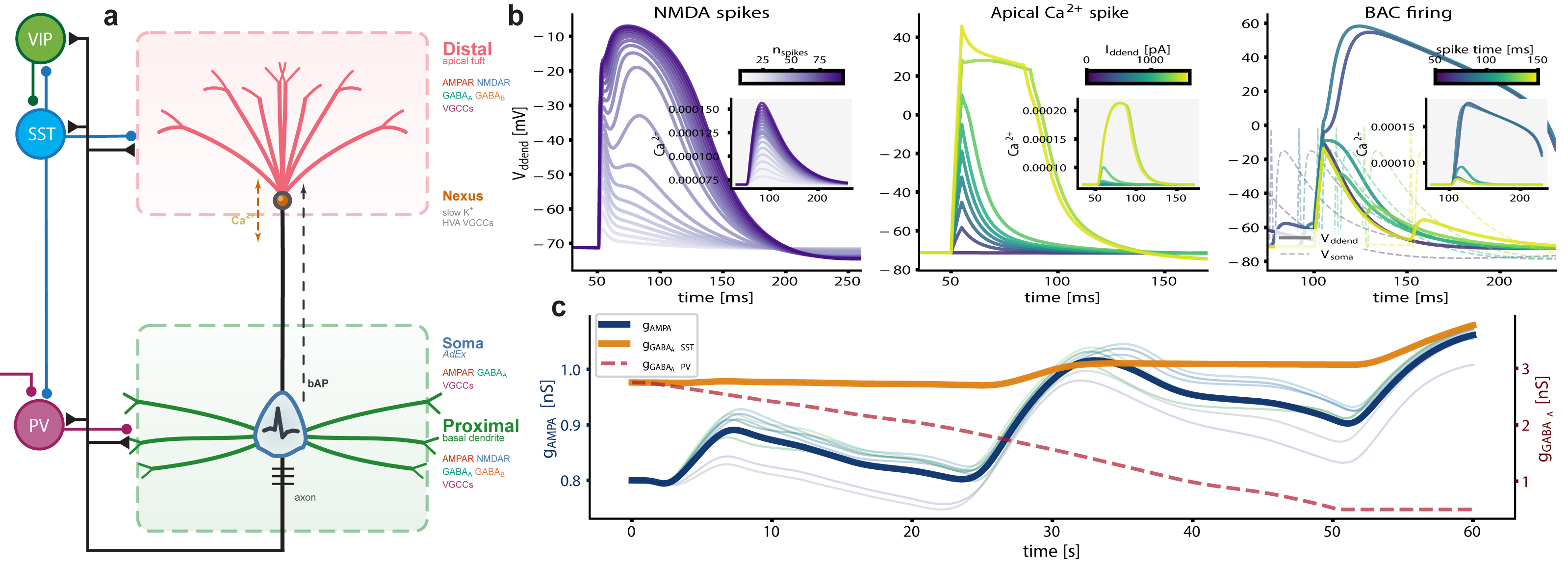 Figure 1. Augmented tripod neuron with compartment-specific electrogenic events and coupled E/I plasticity. (a) Circuit schematic with compartment-resolved receptors and interneuron targeting. (b) NMDA plateaus, apical Ca²⁺ spikes, and BAC firing with dendritic calcium transients (insets). (c) Shared calcium couples excitatory and inhibitory weight dynamics, actively maintaining E/I balance.References
Figure 1. Augmented tripod neuron with compartment-specific electrogenic events and coupled E/I plasticity. (a) Circuit schematic with compartment-resolved receptors and interneuron targeting. (b) NMDA plateaus, apical Ca²⁺ spikes, and BAC firing with dendritic calcium transients (insets). (c) Shared calcium couples excitatory and inhibitory weight dynamics, actively maintaining E/I balance.References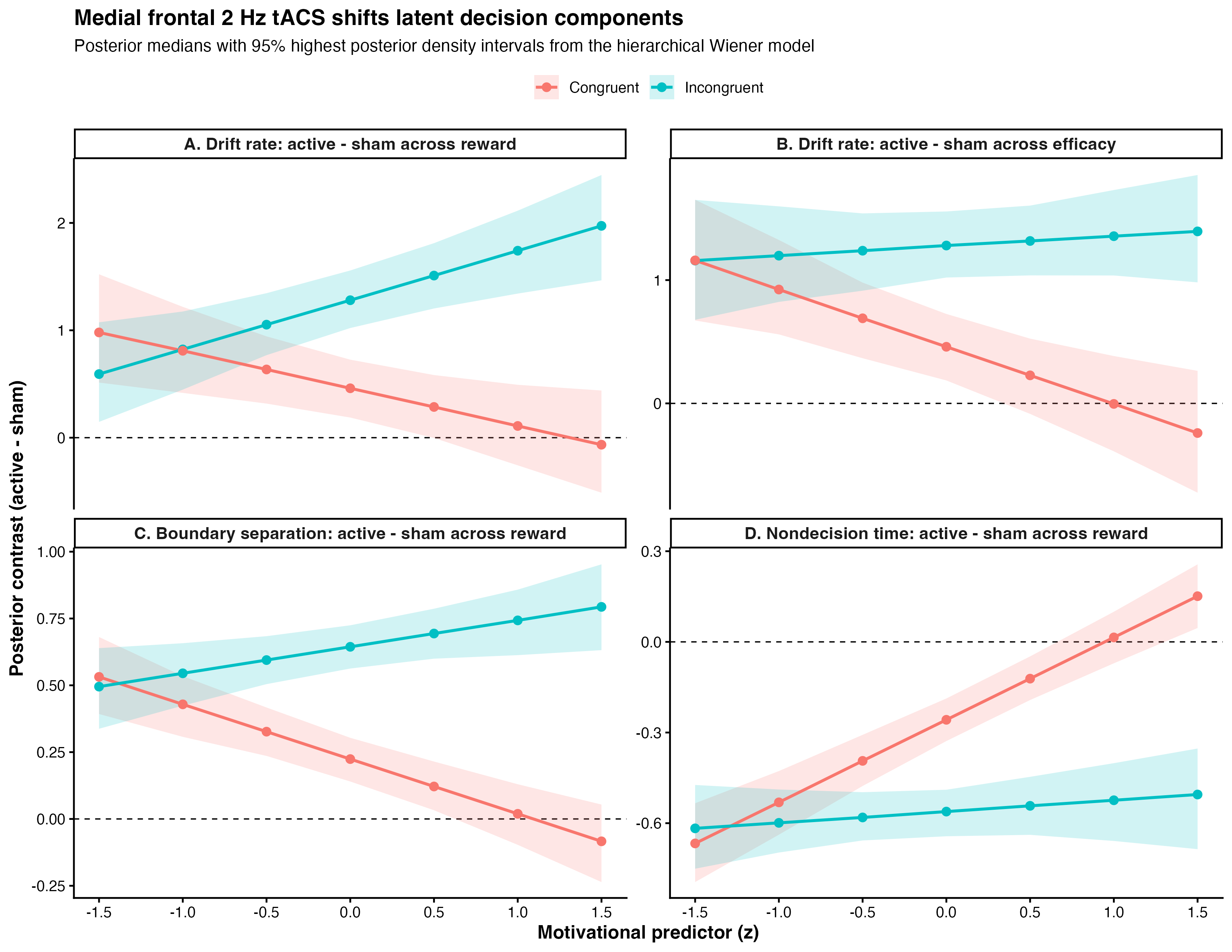 Figure 1. Active minus sham posterior contrasts from the hierarchical Wiener diffusion model. Top panels show drift-rate changes across reward and efficacy, separated by congruency. Bottom panels show boundary-separation and nondecision-time changes across reward. Points indicate posterior medians; ribbons show 95% highest posterior density intervals.
Figure 1. Active minus sham posterior contrasts from the hierarchical Wiener diffusion model. Top panels show drift-rate changes across reward and efficacy, separated by congruency. Bottom panels show boundary-separation and nondecision-time changes across reward. Points indicate posterior medians; ribbons show 95% highest posterior density intervals.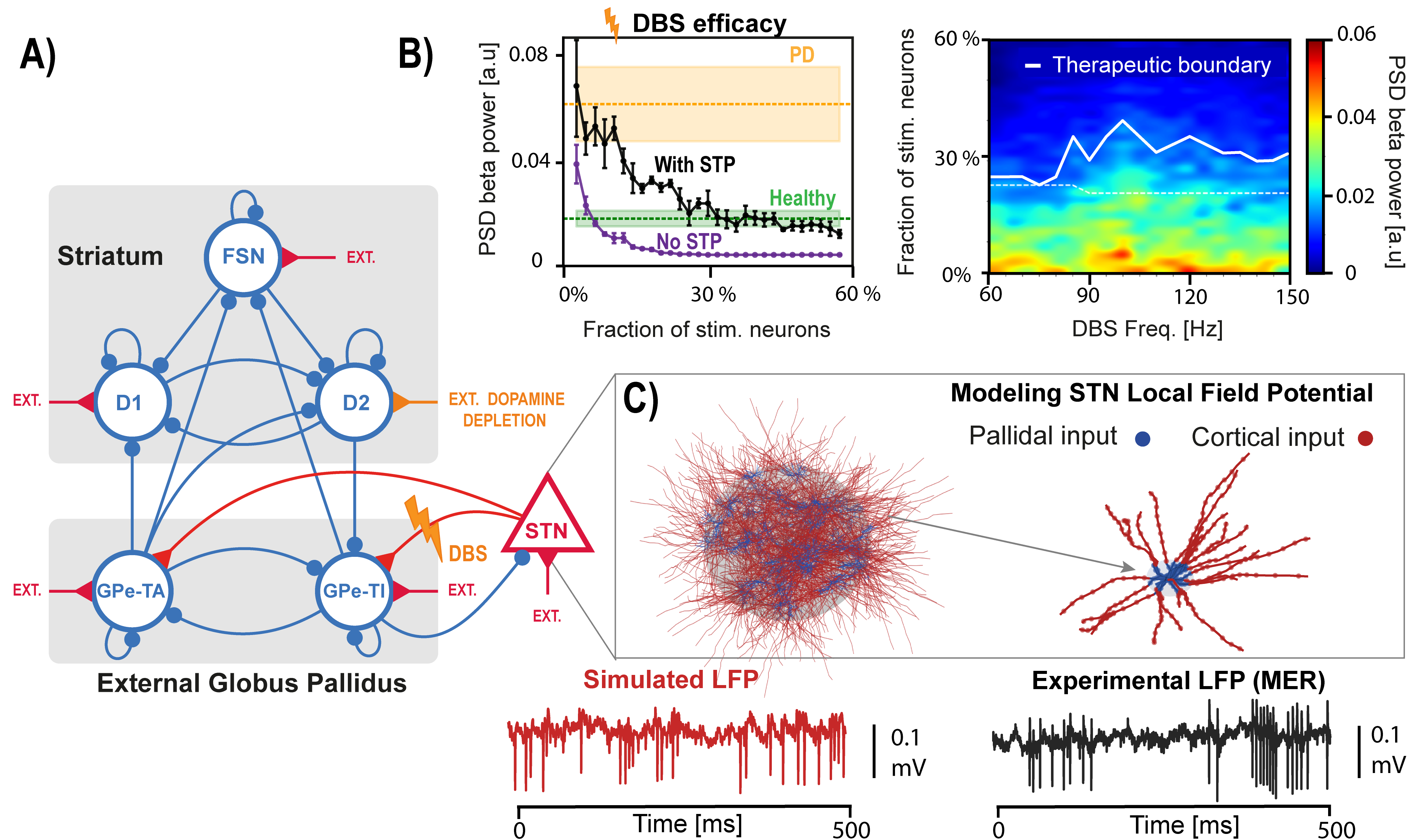 Figure 1. A) Spiking network model of the basal ganglia. B) Left: efficacy of STN DBS as a function of the fraction of stimulated neurons, with and without short-term plasticity (STP). Right: effect of stimulation frequencies on beta spectral power. C) Top: STN population and morphological neuron model receiving cortical and pallidal inputs. Bottom: simulated and recorded local field potentials (LFPs).
Figure 1. A) Spiking network model of the basal ganglia. B) Left: efficacy of STN DBS as a function of the fraction of stimulated neurons, with and without short-term plasticity (STP). Right: effect of stimulation frequencies on beta spectral power. C) Top: STN population and morphological neuron model receiving cortical and pallidal inputs. Bottom: simulated and recorded local field potentials (LFPs).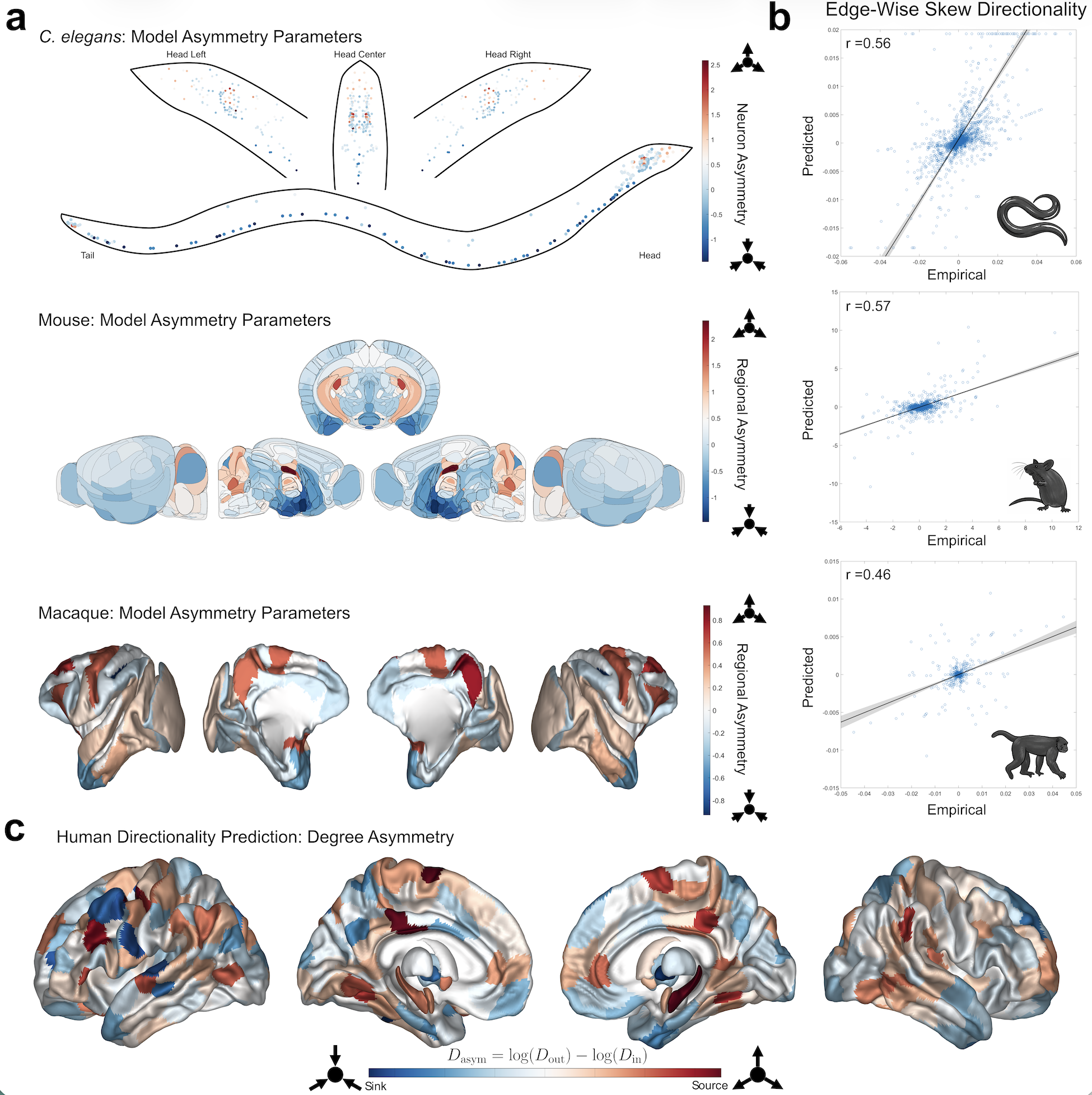 Figure 1. (a) Model-estimated directionality parameters (e^{Ga}) for the three non-human species: C. elegans (top), Mouse (middle), Macaque (bottom). In the C. elegans plot, each dot represents a single neuron. (b) Scatter plots comparing empirical to predicted skew edges with Pearson correlations listed at the top left (all p<10^{-37}). (c) Predicted human overall degree asymmetry for 414 brain regions.
Figure 1. (a) Model-estimated directionality parameters (e^{Ga}) for the three non-human species: C. elegans (top), Mouse (middle), Macaque (bottom). In the C. elegans plot, each dot represents a single neuron. (b) Scatter plots comparing empirical to predicted skew edges with Pearson correlations listed at the top left (all p<10^{-37}). (c) Predicted human overall degree asymmetry for 414 brain regions.
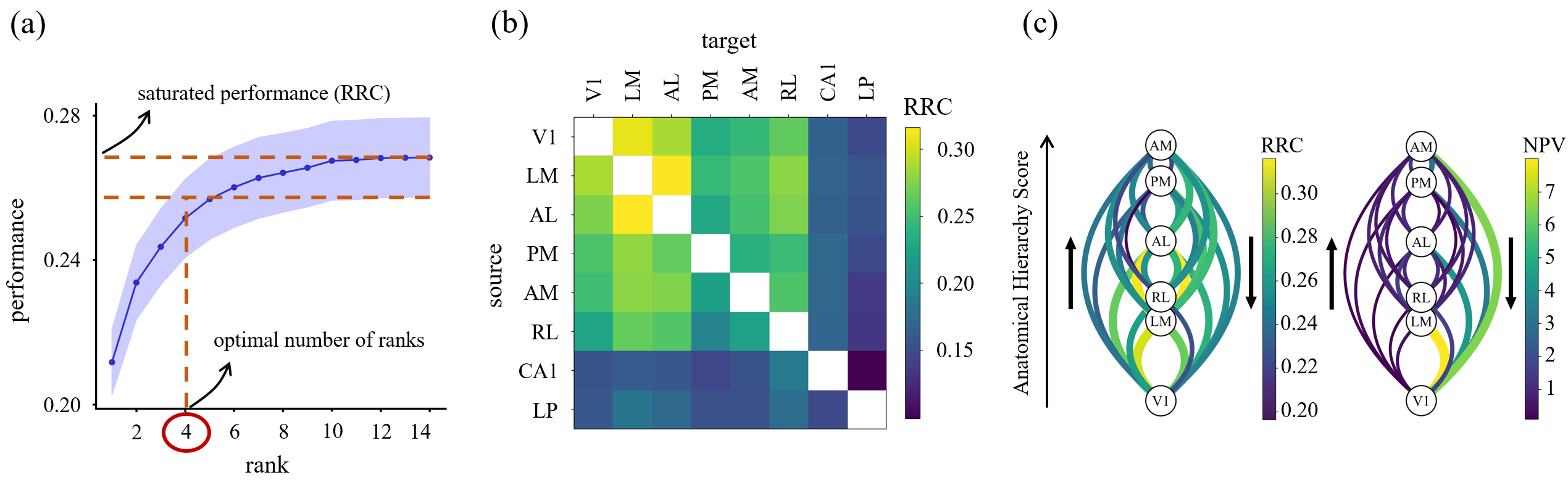 Figure 1. Low-dimensional communication subspaces define RRC. (a) Prediction performance vs. rank; saturation defines optimal number of ranks and RRC. (b) Average RRC across animals; cortical areas are more predictable than subc
Figure 1. Low-dimensional communication subspaces define RRC. (a) Prediction performance vs. rank; saturation defines optimal number of ranks and RRC. (b) Average RRC across animals; cortical areas are more predictable than subc
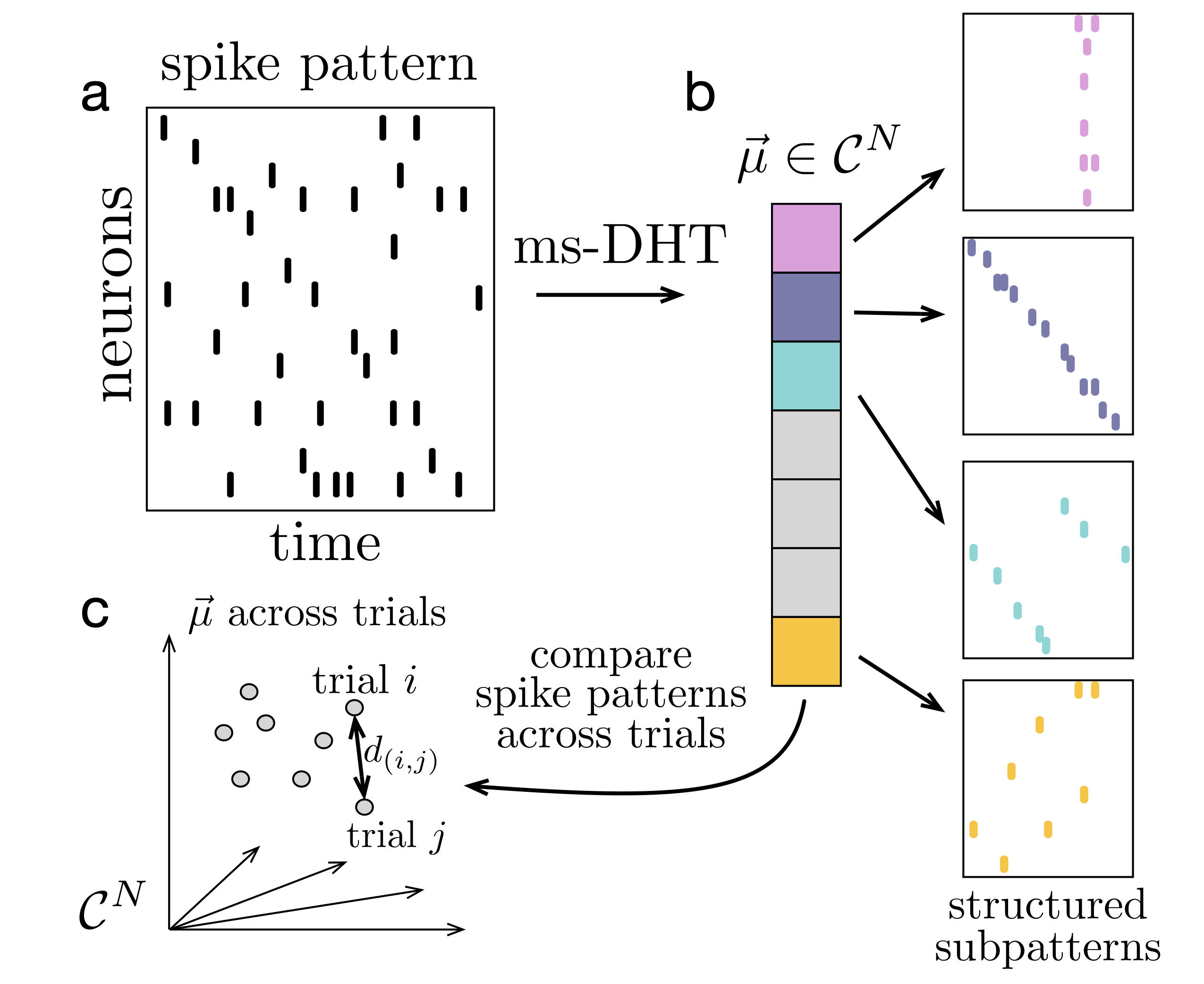 FIgure 1. Decomposing spike patterns. The ms-DHT maps a spike pattern (a) to a unique complex-valued vector (b). Each component encodes the strength (amplitude) and timing (phase) of a basis sub-pattern. (c) Distances between spike patterns reduce to Euclidean distances between ms-DHT outputs, which are invariant to neuron order, ensuring behaviourally relevant clusters do not depend on neuron order.
FIgure 1. Decomposing spike patterns. The ms-DHT maps a spike pattern (a) to a unique complex-valued vector (b). Each component encodes the strength (amplitude) and timing (phase) of a basis sub-pattern. (c) Distances between spike patterns reduce to Euclidean distances between ms-DHT outputs, which are invariant to neuron order, ensuring behaviourally relevant clusters do not depend on neuron order. Figure 1. The CL-1 device is scalable desktop device compatible with standard server racks that allows real-time closed-loop interactions with neural cells via an MEA reader. The CL-1 has onboard hardware that interprets simple code via a Python API to allow rapid code development and experimental iterations coupled with a closed-loop perfusion circuit to automatically adjusts gas levels and temperature to
Figure 1. The CL-1 device is scalable desktop device compatible with standard server racks that allows real-time closed-loop interactions with neural cells via an MEA reader. The CL-1 has onboard hardware that interprets simple code via a Python API to allow rapid code development and experimental iterations coupled with a closed-loop perfusion circuit to automatically adjusts gas levels and temperature to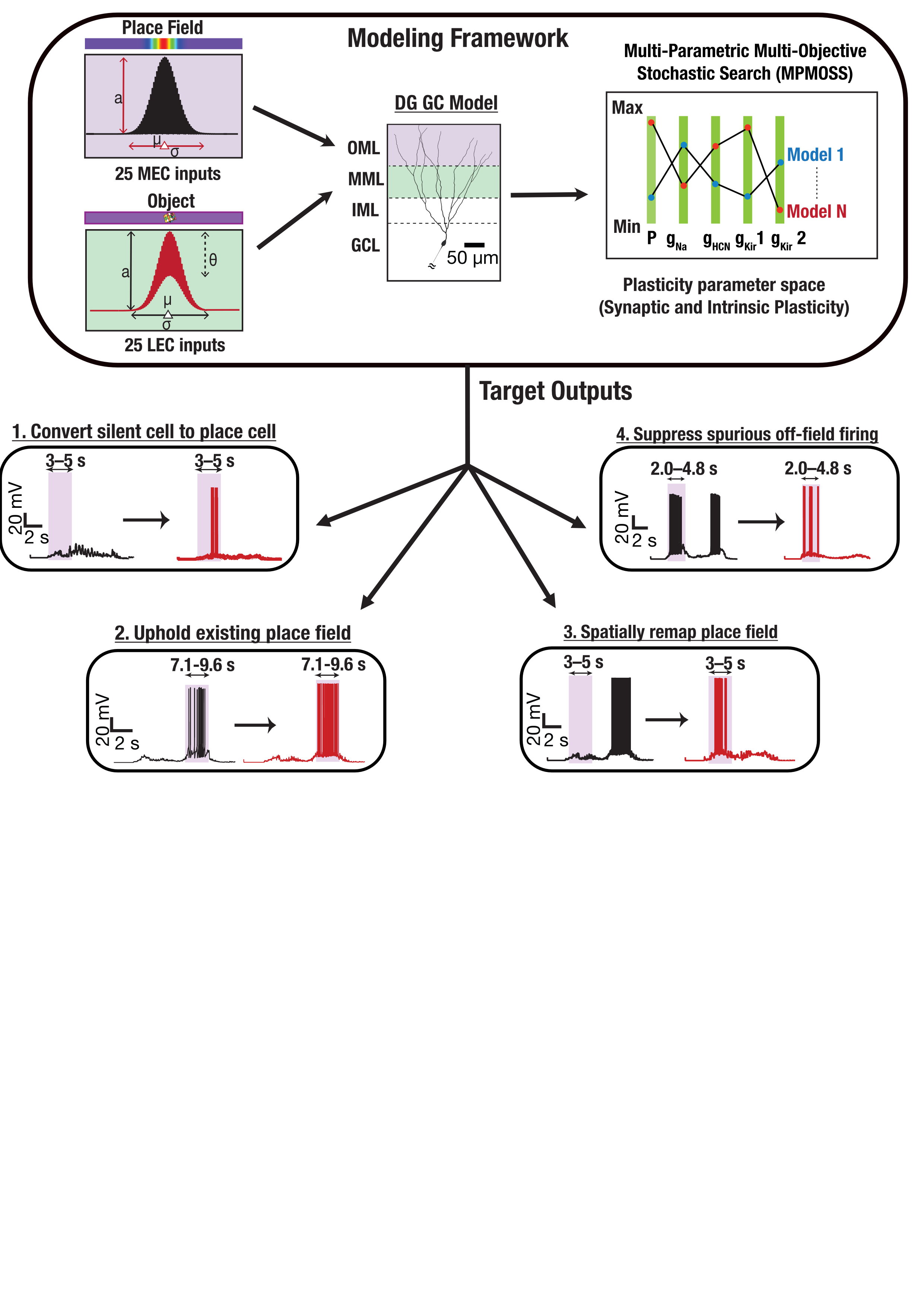 FIgure 1. Medial and lateral entorhinal cortex inputs impinge on a DG granule cell. Disparate combinations of synaptic and intrinsic plasticity (NaP, HCN, Kir channels) achieved one of four targets: convert silent cell to place cell, uphold existing place field, remap, or suppress spurious firing. Our results show that robust and flexible spatial tuning is achievable through plasticity degeneracy.References
FIgure 1. Medial and lateral entorhinal cortex inputs impinge on a DG granule cell. Disparate combinations of synaptic and intrinsic plasticity (NaP, HCN, Kir channels) achieved one of four targets: convert silent cell to place cell, uphold existing place field, remap, or suppress spurious firing. Our results show that robust and flexible spatial tuning is achievable through plasticity degeneracy.References



