Dhuruva Priyan Gowri Mariyappan
*1,2,3, Nghi V Nguyen
2,3,4, Giuseppe Chindemi
5, András Ecker
6, Sabrina Tazerart
2,4, James Isbister
7, Darshan Mandge
7, Diana E. Mitchell
2,4, Michael W Reimann
7, Roberto Araya
4,2, Eilif B Muller
2,3,41 Department of Computer Science and Operations Research, Faculty of Arts and Science, Université de Montréal, Montréal, Canada
2 Centre de Recherche Azrieli du CHU Sainte-Justine, Montréal, Canada
3 Mila Quebec AI Institute, Montréal, Canada
4 Department of Neurosciences, Faculty of Medicine, Université de Montréal, Montréal, Canada
5 ETH AI Center, Zurich, Switzerland
6 Cytocast Hungary Kft, Budapest, Hungary
7 Open Brain Institute, Lausanne, Switzerland
*Email:
[email protected]Introduction
Cortical representations are thought to arise from stable network motifs formed by neuronal assemblies, with synaptic plasticity between pyramidal cells (PCs) playing a central role in their formation, competition, and maintenance. While rules governing such synaptic changes have been well characterized in slice conditions, their implications for learning in awake behaving animals remain an active area of research. Here we use biophysically detailed simulations with calibrated ion channels, background synaptic activity, and calcium-based plasticity rules to investigate mechanisms enabling reliable plasticity in-vivo. We find that spatially clustered activation and burst firing offer robust pathways for LTP under physiological conditions.MethodsWe used biophysically detailed simulations of a large-scale in-silico cortical microcircuit of rat somatosensory cortex with a calcium-based plasticity model capturing LTP and Long-Term Depression (LTD) as a function of integrated postsynaptic calcium. We parameterized voltage-gated Na⁺, K⁺, Ca²⁺, and Bk channels throughout the dendritic tree based on experimental data. To reproduce the high-conductance state of awake cortex, we incorporated stochastic background activity using Dendritic Extra-Excitatory Synapses (DEES) at 1.1 synapses/μm. We investigated clustered plasticity in L2/3 PC and L5-TTPC basal and apical dendrites under both in-vitro and in-vivo-like extracellular calcium concentrations.ResultsSynchronous activation of ≥11 clustered synapses generates dendritic plateau potentials (≥100 ms) that induce LTP in distal basal dendrites (Fig. 1). We identify a synapse-pairing tradeoff, where dendrites effectively trade the number of co-activated synapses for pairing repetitions: 16-synapse clusters achieve one-shot learning, while 8-synapse clusters require 3+ pairings. This tradeoff exhibits spatial gradients: distal dendrites achieve LTP independent of backpropagating action potentials, while proximal clusters require spike-timing coincidence. When multiple basal clusters coactivate, summated depolarizations trigger somatic bursts; both presynaptic and postsynaptic bursts drive robust LTP with all-or-none threshold dynamics.
DiscussionThese findings establish multiple plasticity mechanisms within a single neuron—spatial clustering, location-dependent learning modes, and dual burst pathways—providing testable predictions for how cortical circuits implement flexible, hierarchical learning. Distal dendrites enable unsupervised learning via cluster-based LTP independent of bAPs, while proximal regions implement supervised learning requiring spike-timing coincidence. Apical dendrites receiving top-down signals can generate bursts or couple with somatic spikes via backpropagation-activated calcium (BAC) firing, a substrate for top-down plasticity modulation. These mechanisms reveal how dendrites trade synapse number for pairing repetitions under noisy physiological conditions.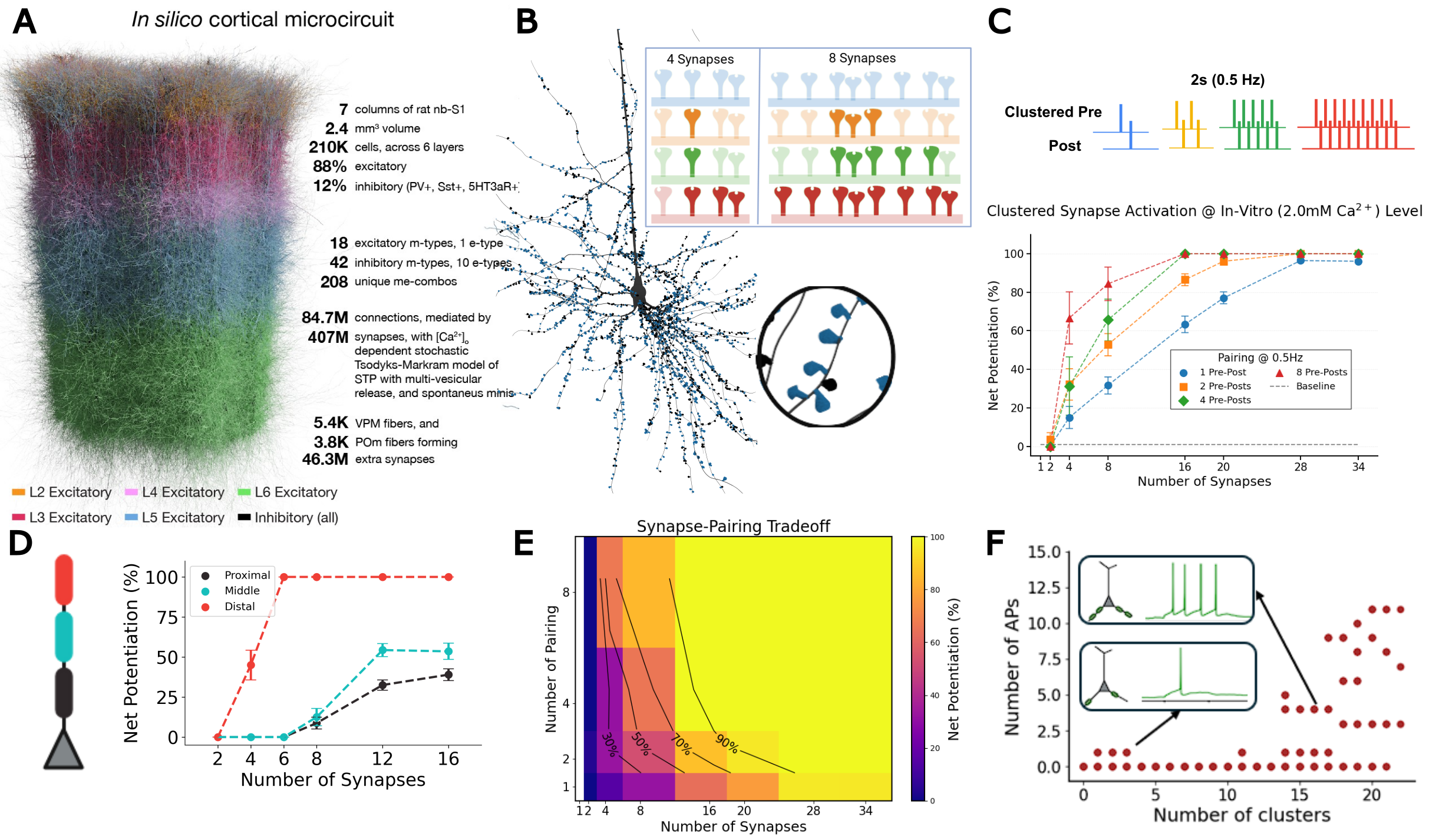 Figure 1. A, In silico cortical microcircuit. B, L5-TTPC with magnified cluster showing plasticity for 4 vs 8 co-active synapses. C, Clustered pre-post pairing (0.5 Hz); net potentiation vs synapse number. D, Spatial learning gradient E, Synapse-pairing tradeoff heatmap. F, Basal cluster coactivation triggers somatic burst.
Figure 1. A, In silico cortical microcircuit. B, L5-TTPC with magnified cluster showing plasticity for 4 vs 8 co-active synapses. C, Clustered pre-post pairing (0.5 Hz); net potentiation vs synapse number. D, Spatial learning gradient E, Synapse-pairing tradeoff heatmap. F, Basal cluster coactivation triggers somatic burst.
References1. Chindemi, G., Abdellah, M., Amsalem, O., Benavides-Piccione, R., Delattre, V., Doron, M., Ecker, A., Jaquier, A. T., King, J., Kumbhar, P., Monney, C., Perin, R., Rössert, C., Tuncel, A. M., Van Geit, W., DeFelipe, J., Graupner, M., Segev, I., Markram, H., & Muller, E. B. (2022). A calcium-based plasticity model for predicting long-term potentiation and depression in the neocortex.
2. Ecker, A., Egas Santander, D., Abdellah, M., Alonso, J. B., Bolaños-Puchet, S., Chindemi, G., Gowri Mariyappan, D. P., Isbister, J. B., Ki
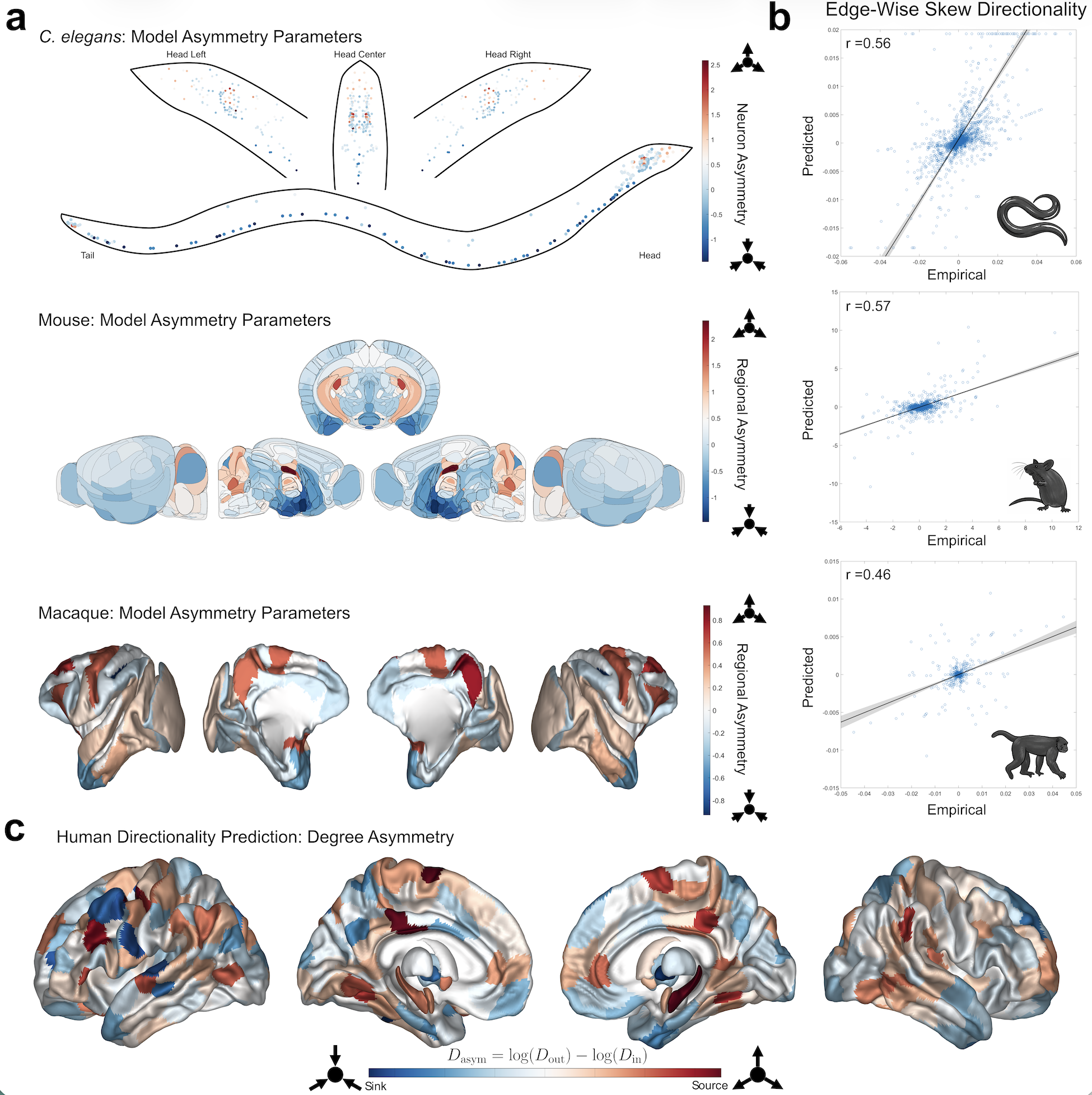 Figure 1. (a) Model-estimated directionality parameters (e^{Ga}) for the three non-human species: C. elegans (top), Mouse (middle), Macaque (bottom). In the C. elegans plot, each dot represents a single neuron. (b) Scatter plots comparing empirical to predicted skew edges with Pearson correlations listed at the top left (all p<10^{-37}). (c) Predicted human overall degree asymmetry for 414 brain regions.
Figure 1. (a) Model-estimated directionality parameters (e^{Ga}) for the three non-human species: C. elegans (top), Mouse (middle), Macaque (bottom). In the C. elegans plot, each dot represents a single neuron. (b) Scatter plots comparing empirical to predicted skew edges with Pearson correlations listed at the top left (all p<10^{-37}). (c) Predicted human overall degree asymmetry for 414 brain regions.
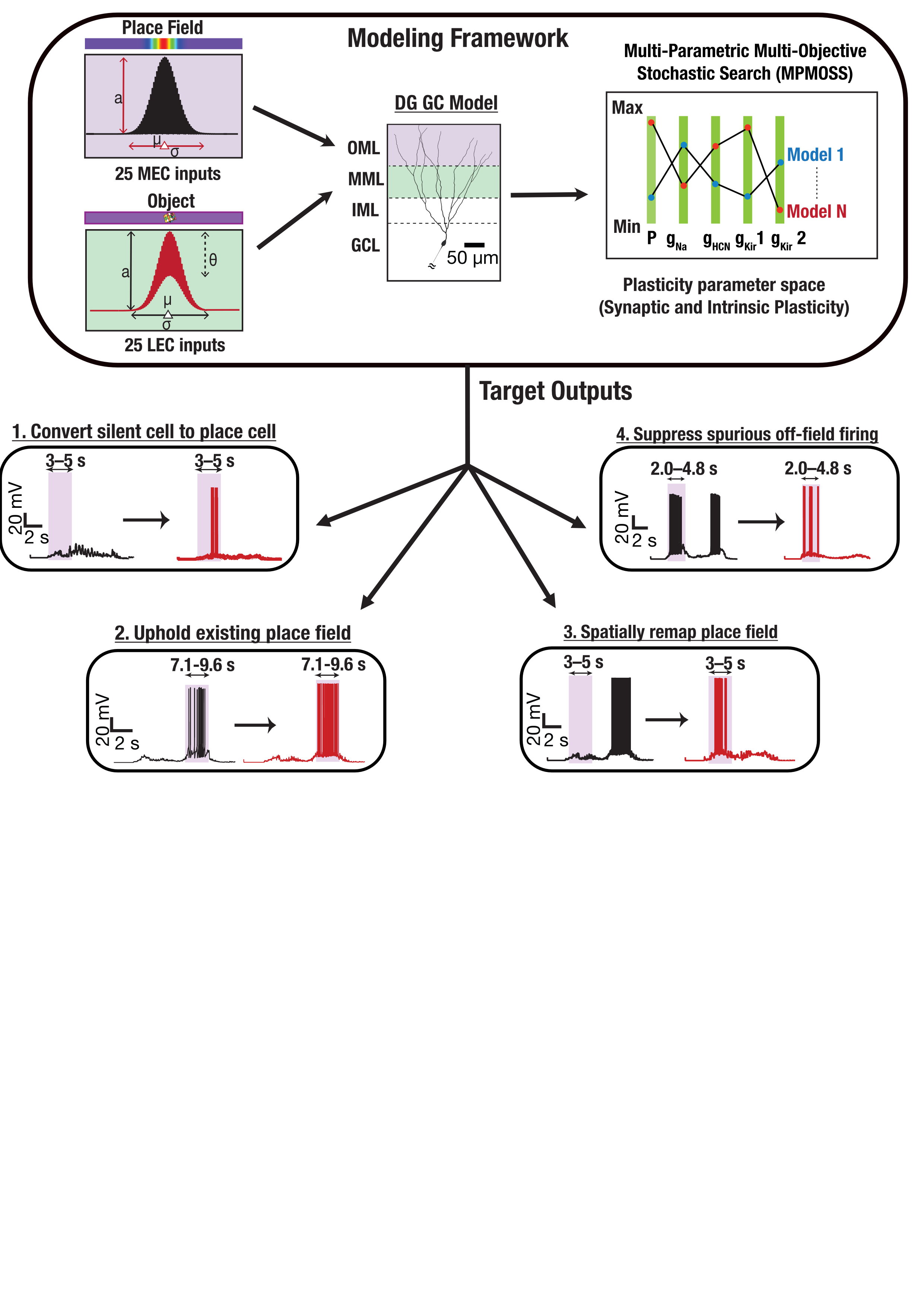 FIgure 1. Medial and lateral entorhinal cortex inputs impinge on a DG granule cell. Disparate combinations of synaptic and intrinsic plasticity (NaP, HCN, Kir channels) achieved one of four targets: convert silent cell to place cell, uphold existing place field, remap, or suppress spurious firing. Our results show that robust and flexible spatial tuning is achievable through plasticity degeneracy.References
FIgure 1. Medial and lateral entorhinal cortex inputs impinge on a DG granule cell. Disparate combinations of synaptic and intrinsic plasticity (NaP, HCN, Kir channels) achieved one of four targets: convert silent cell to place cell, uphold existing place field, remap, or suppress spurious firing. Our results show that robust and flexible spatial tuning is achievable through plasticity degeneracy.References



