IntroductionA comprehensive behavioural decision can be made based on a preliminary aesthetic assessment that harmonizes a variety of factors. Subjective prediction can cover at least 20-30% of brain regions, from sensory and motor areas to the cerebral cortex [1]. To study such processes, neuroaesthetic tools are being developed that combine computational rating models with neurophysiological methods [2].
In our work, we analysed how an aesthetic assessment of a location affects the choice of route for a traveller through natural landscapes. We have shown that computational neuroaesthetics, supplemented by spatial trajectory analysis, can be used to identify significant landscape factors and to predict attractiveness of tourist destinations.
MethodsOur goal was to identify indicators that determine the aesthetic appeal of natural attractions. We calculated topological and visual characteristics for three different tourist sites near Bishkek, Kyrgyzstan, with different Google Maps ratings and different frequencies of mentions on Internet (FM) (Figure 1):
A) topological features of routes and average densities of tourist flows (attendance) based on GPS tracks of activity tracking service (https://www.strava.com);
B) visibility pools and visual perception trails, softness of relief lines and spot colour balance based on remote sensing data.
The analysis was performed using QGIS spatial tools (https://qgis.org).
ResultsWe performed calculations for three tourist locations and compared the results with 6 non-tourist locations near Bishkek. Then we identified topological and visual indicators that differed by at least 25% between tourist sites and other locations.
Based on our analysis, we determined the "comfortable" properties of the locations, such as: visual openness of space, softness of lines, neutral natural colours.
Similar quality metrics of visual walkability perception in urban pedestrians have been found by Li Y et al. using panoramic street view images, virtual reality, and deep learning [3].
The applied methods and identified indicators can be used in machine learning tasks of artificial neural networks to detect high-rated tourist areas.
DiscussionWhen receiving information from different sensory systems, the brain processes it, forming a complex response to multi-layered data sets.
Aesthetic ratings of natural scenes can correlate with empirical data on visual comfort, while aesthetic preferences may be driven by optimization of decision-making processes in favour of lower-energy states of brain activity [4].
This is consistent with tourists\' reviews of natural attractions near Bishkek as "soothing" and "relaxing" locations.
In future studies, we plan to continue studying the influence of aesthetic characteristics of complex spatial stimuli on route selection, including comparing data on brain activity and data on movement trajectories.
 Figure 1. A1, A2: GPS tracks and photo of the location "Ala-Archa" (FM – 1.6 million, attendance 215 people/hour, rating 4.7), B1, B2: GPS tracks and photo of the location "Sky Bridge" (FM – 39.8 thousand, attendance 36 people/hour, rating 4.5). C1, C2: GPS tracks and a photo of the location "Raspberry Gorge" (FM 2.9 thousand, attendance 4 people/hour, rating 4.0).
Figure 1. A1, A2: GPS tracks and photo of the location "Ala-Archa" (FM – 1.6 million, attendance 215 people/hour, rating 4.7), B1, B2: GPS tracks and photo of the location "Sky Bridge" (FM – 39.8 thousand, attendance 36 people/hour, rating 4.5). C1, C2: GPS tracks and a photo of the location "Raspberry Gorge" (FM 2.9 thousand, attendance 4 people/hour, rating 4.0).
References1. Findling, C., et al. (2025). Brain-wide representations of prior information in mouse decision-making. Nature, 645, 192–200. https://doi.org/10.1038/s41586-025-09226-1
2. Li, R., & Zhang, J. (2020). Review of computational neuroaesthetics: bridging the gap between neuroaesthetics and computer science. Brain Informatics, 7, 16. https://doi.org/10.1186/s40708-020-00118-w
3. Li, Y., et al. (2022). Measuring visual walkability perception using panoramic street view images, virtual reality, and deep learning. Sustain Cities Soc, 86, 104140. https://doi.org/10.1016/j.scs.2022.104140
4. Tang, Y., et al. (2025). Less is more: Aesthetic liking is inversely related to metabolic expense by the visual system. PNAS Nexus, 4, pgaf347. https://doi.org/10.1093/pnasnexus/pgaf347
AcknowledgementWe gratefully acknowledge the applications and tools provided by QGIS. This platform is constantly evolving, expanding the possibilities of spatial analysis for all fields of knowledge.
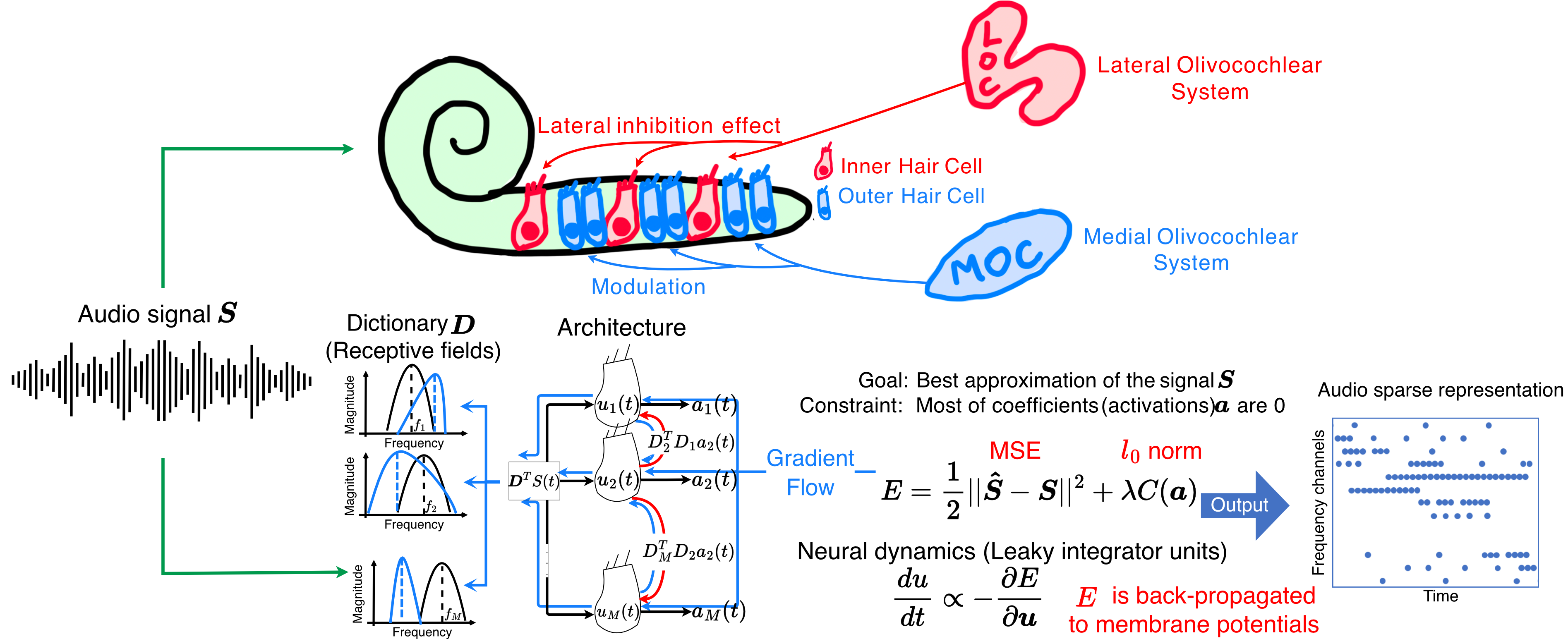 Figure 1. Overview of the ALCA-CF front-end. Each filter is represented by a neuron with a receptive field. Red arrows are lateral inhibition synapses and blue arrows are the feedback that adapts each neuron's receptive field. Lateral inhibition weights are the correlation between pre- and post-synaptic neuron receptive fields. Neuron activations ai are represented by blue dots in the time-frequency output.
Figure 1. Overview of the ALCA-CF front-end. Each filter is represented by a neuron with a receptive field. Red arrows are lateral inhibition synapses and blue arrows are the feedback that adapts each neuron's receptive field. Lateral inhibition weights are the correlation between pre- and post-synaptic neuron receptive fields. Neuron activations ai are represented by blue dots in the time-frequency output.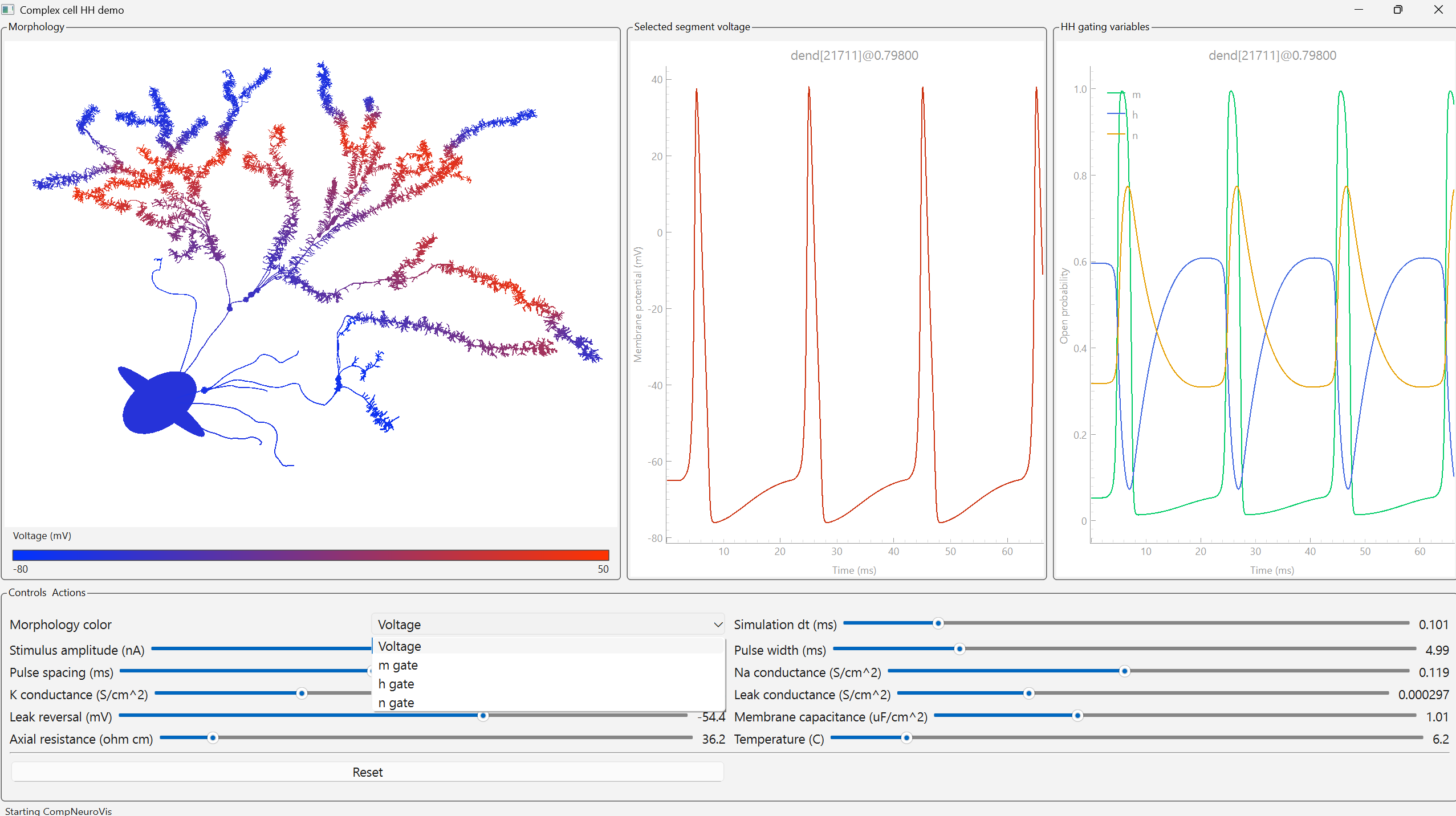 Figure 1. A CompNeuroVis application built around a compartmental NEURON model of a reconstructed cell with Hodgkin-Huxley dynamics. The morphology is color-coded by a selected variable, here membrane voltage. For a selected segment, linked plots show membrane voltage and the gating variables m, h, and n. A dropdown sets the mapped variable and sliders adjust stimulus and biophysical parameters.
Figure 1. A CompNeuroVis application built around a compartmental NEURON model of a reconstructed cell with Hodgkin-Huxley dynamics. The morphology is color-coded by a selected variable, here membrane voltage. For a selected segment, linked plots show membrane voltage and the gating variables m, h, and n. A dropdown sets the mapped variable and sliders adjust stimulus and biophysical parameters.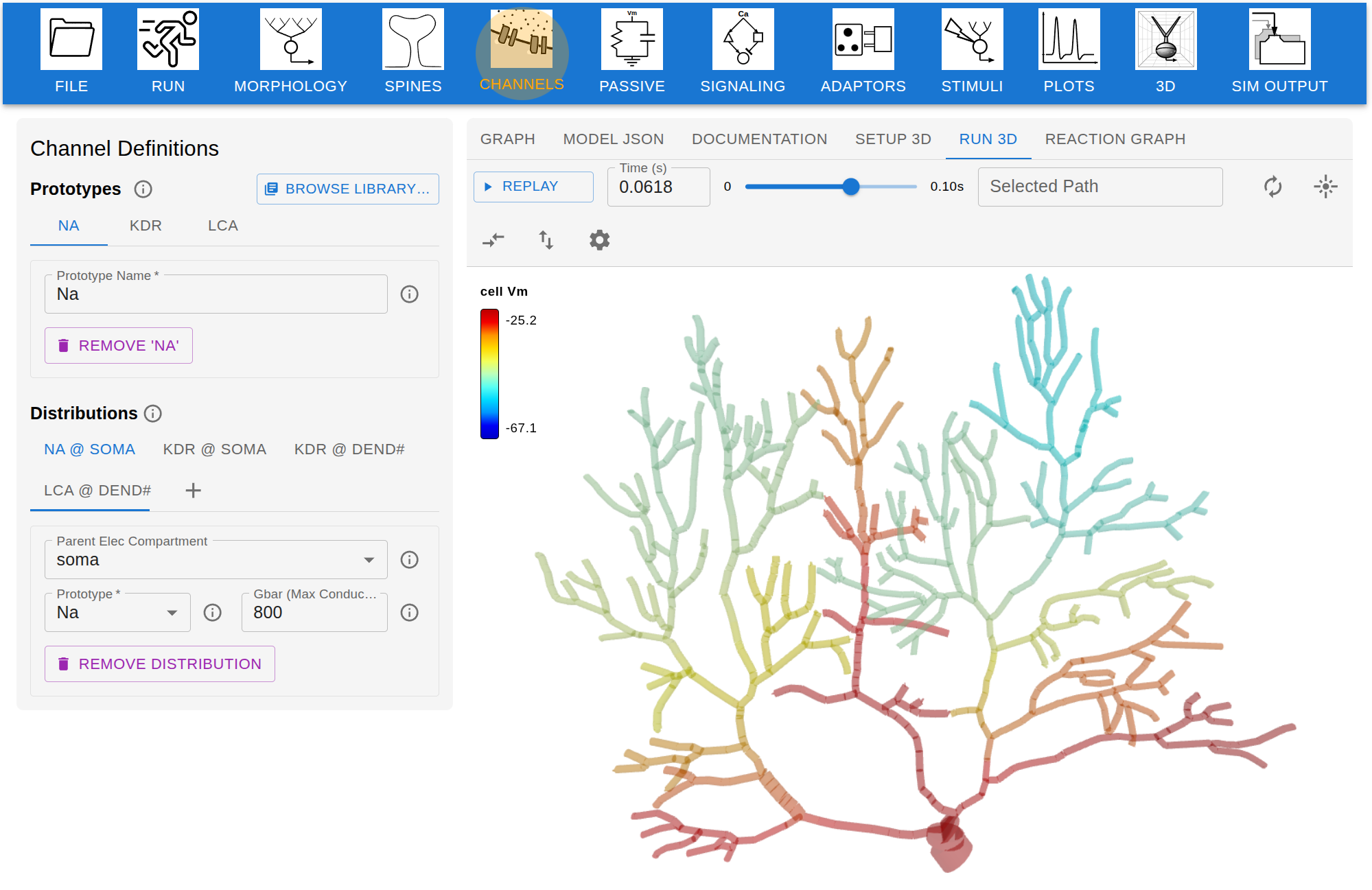 Figure 1. Screenshot of Jardesigner GUI. The menu options are in the blue bar above, and the Channel Menu box is open to the left. A snapshot of the current simulation is presented in the 3D display to the right.
Figure 1. Screenshot of Jardesigner GUI. The menu options are in the blue bar above, and the Channel Menu box is open to the left. A snapshot of the current simulation is presented in the 3D display to the right.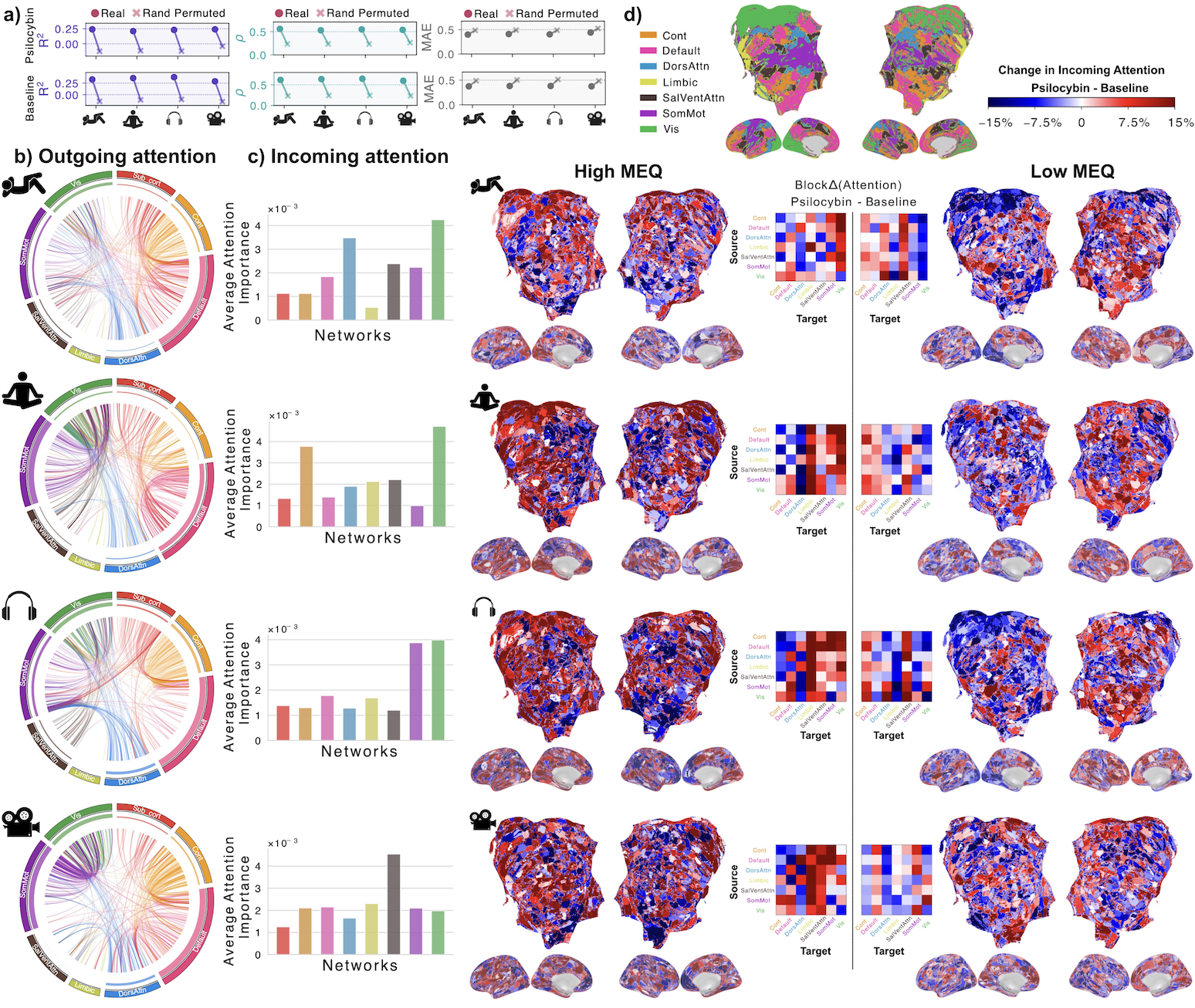 Figure 1. BrainSymphony reconstruction and attention-based reorganization. (a) Paired dots: real vs permuted ROI series across conditions; higher R²,r, lower MAE. (b) Circos: Admin–Baseline attention Δ (top 500 edges) colored by source network; inner track = total outgoing. (c) Network-mean incoming attention Δ. (d) High vs Low MEQ receptive-attention maps plus inter-network Δ matrices.
Figure 1. BrainSymphony reconstruction and attention-based reorganization. (a) Paired dots: real vs permuted ROI series across conditions; higher R²,r, lower MAE. (b) Circos: Admin–Baseline attention Δ (top 500 edges) colored by source network; inner track = total outgoing. (c) Network-mean incoming attention Δ. (d) High vs Low MEQ receptive-attention maps plus inter-network Δ matrices.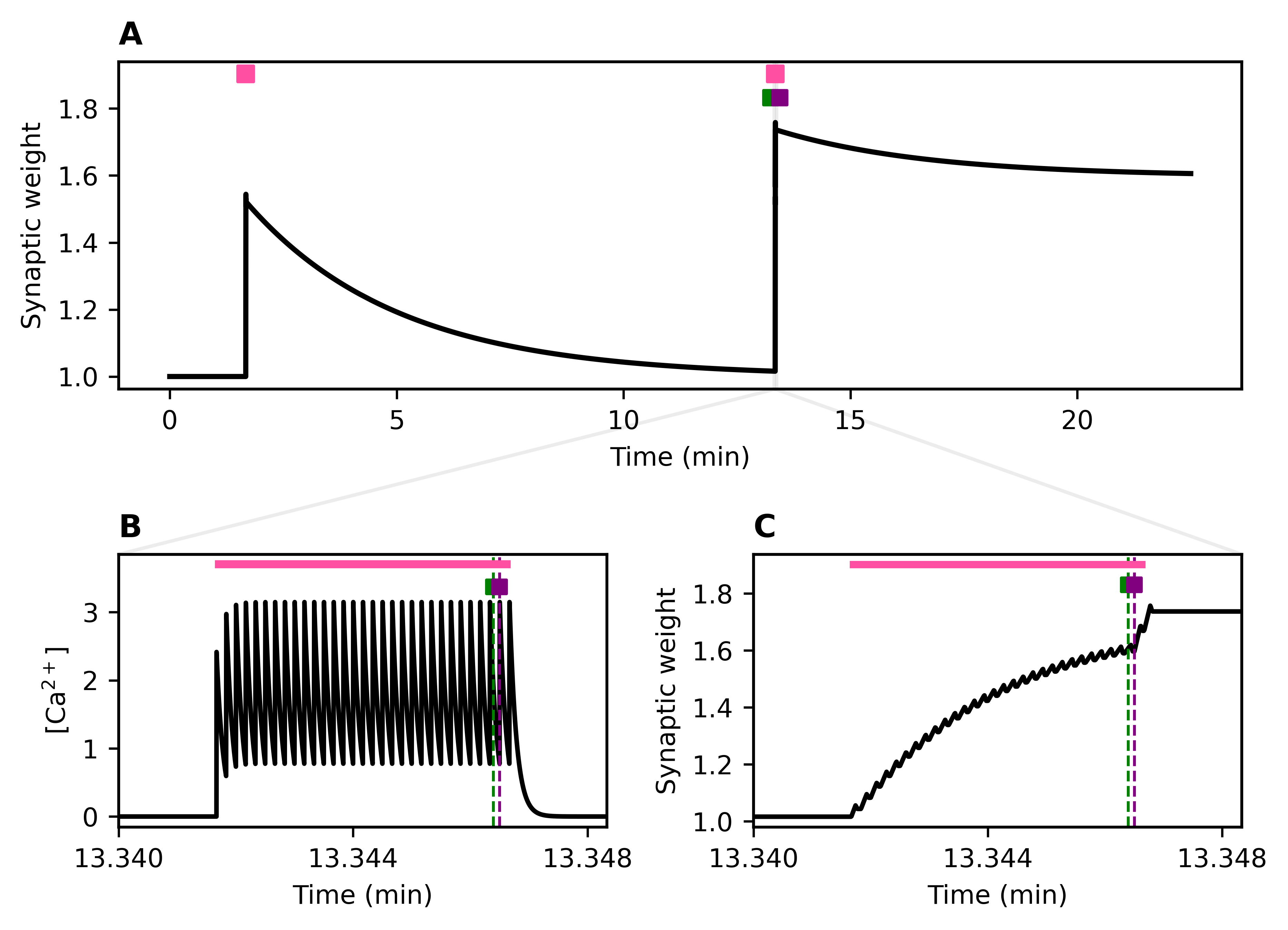 Figure 1. Stochastic activity-driven ER entry and stabilization preserve calcium-dependent plasticity over long timescales. (A) Identical high-frequency pre/post spike trains (pink) produce synaptic weight changes; the first stimulation fails to trigger ER entry, while the second induces ER entry (green) followed by stabilization (purple). (B–C) Insets show calcium (left) and weight (right) dynamicsReferences
Figure 1. Stochastic activity-driven ER entry and stabilization preserve calcium-dependent plasticity over long timescales. (A) Identical high-frequency pre/post spike trains (pink) produce synaptic weight changes; the first stimulation fails to trigger ER entry, while the second induces ER entry (green) followed by stabilization (purple). (B–C) Insets show calcium (left) and weight (right) dynamicsReferences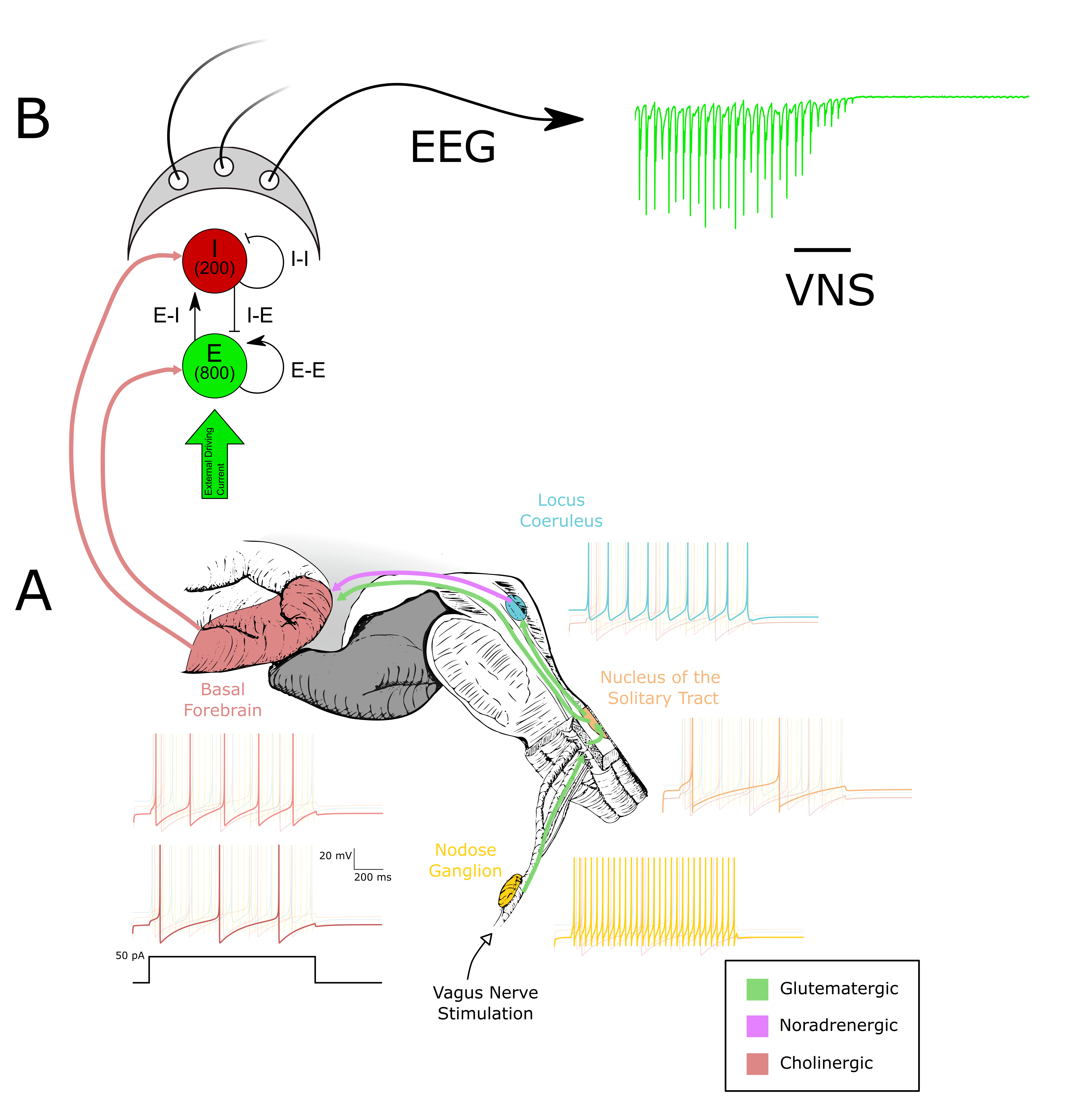 Figure 1. (A) The circuitry between the vagus nerve and the basal forebrain is modeled using quadratic integrate-and-fire (Izhikevich) neurons fitted to experimental data. Representative voltage responses to a 50 pA injection are shown. (B) The output of the basal forebrain in (A) is used to modulate ACh-sensitive K+ channels in an E–I network. Synaptic activity within this network is used to compute an EEGReferences
Figure 1. (A) The circuitry between the vagus nerve and the basal forebrain is modeled using quadratic integrate-and-fire (Izhikevich) neurons fitted to experimental data. Representative voltage responses to a 50 pA injection are shown. (B) The output of the basal forebrain in (A) is used to modulate ACh-sensitive K+ channels in an E–I network. Synaptic activity within this network is used to compute an EEGReferences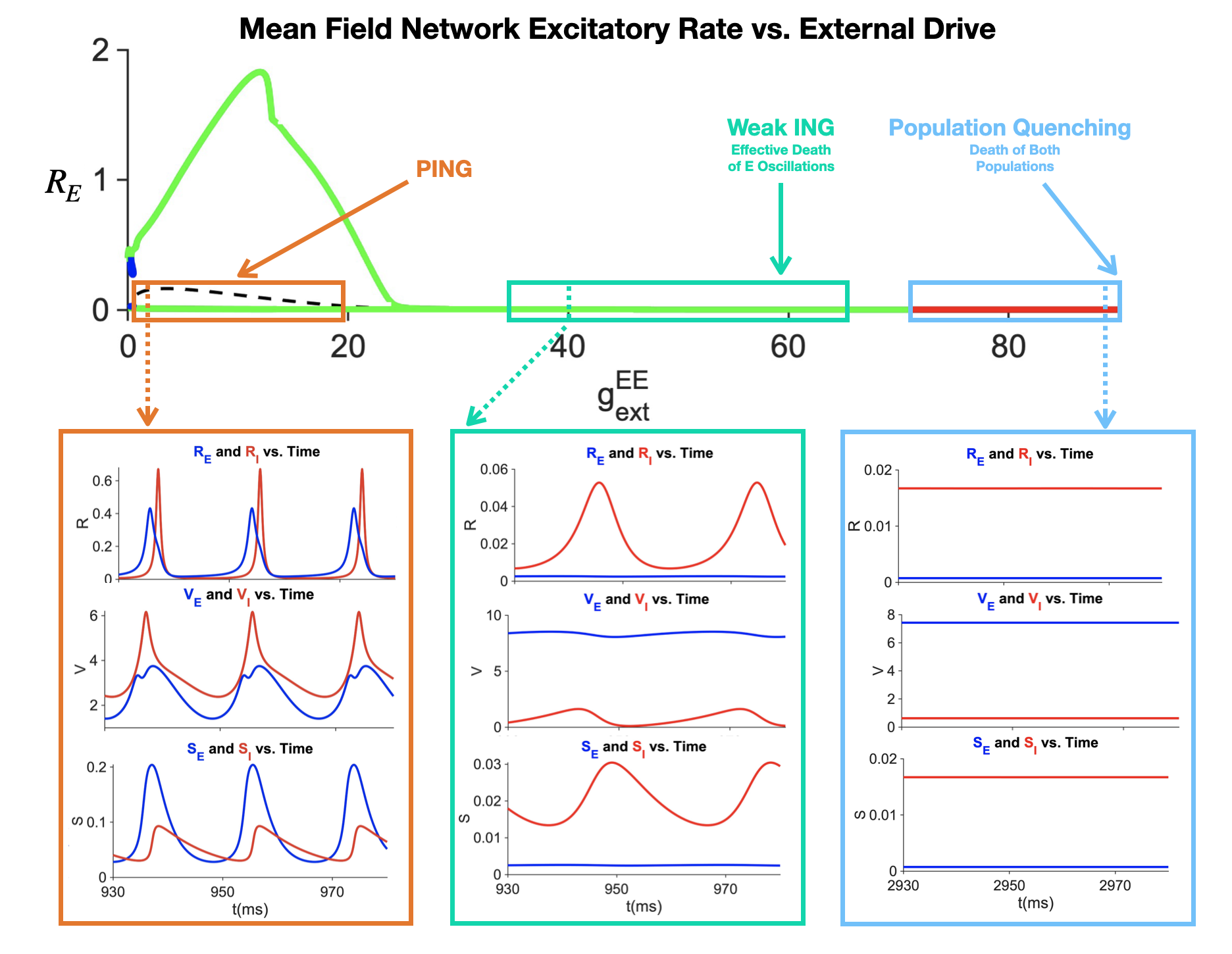 Figure 1. E-I Network dynamics as a function of external excitatory conductance drive in high input regime
Figure 1. E-I Network dynamics as a function of external excitatory conductance drive in high input regime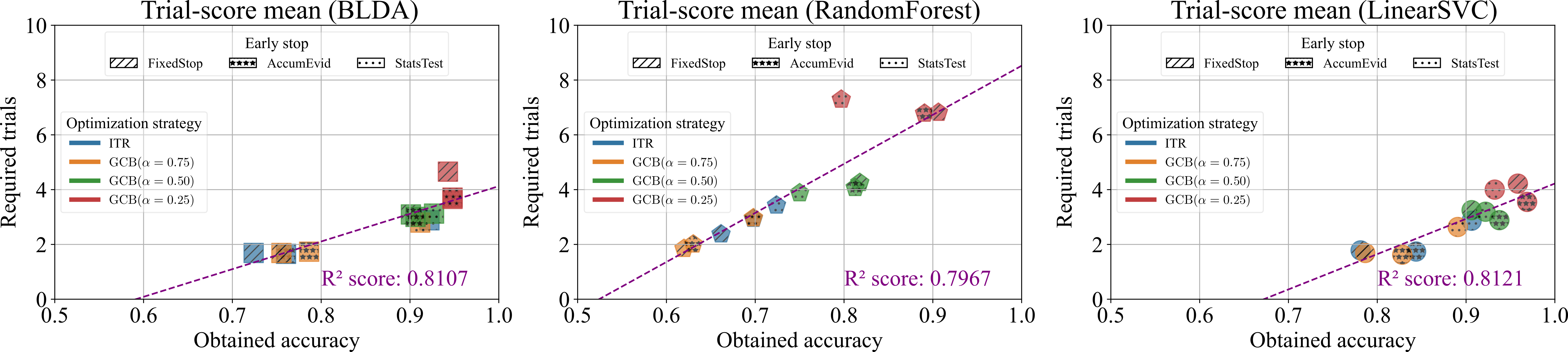 Figure 1. Required trials and obtained accuracies across classifiers, early-stopping strategies, and optimization methods for Hoffmann et. al. Rapid Serial Visual Presentation dataset [1]. Points show mean performance under leave-one-session-out validation. The fitted line illustrates the speed–accuracy trade-off.
Figure 1. Required trials and obtained accuracies across classifiers, early-stopping strategies, and optimization methods for Hoffmann et. al. Rapid Serial Visual Presentation dataset [1]. Points show mean performance under leave-one-session-out validation. The fitted line illustrates the speed–accuracy trade-off.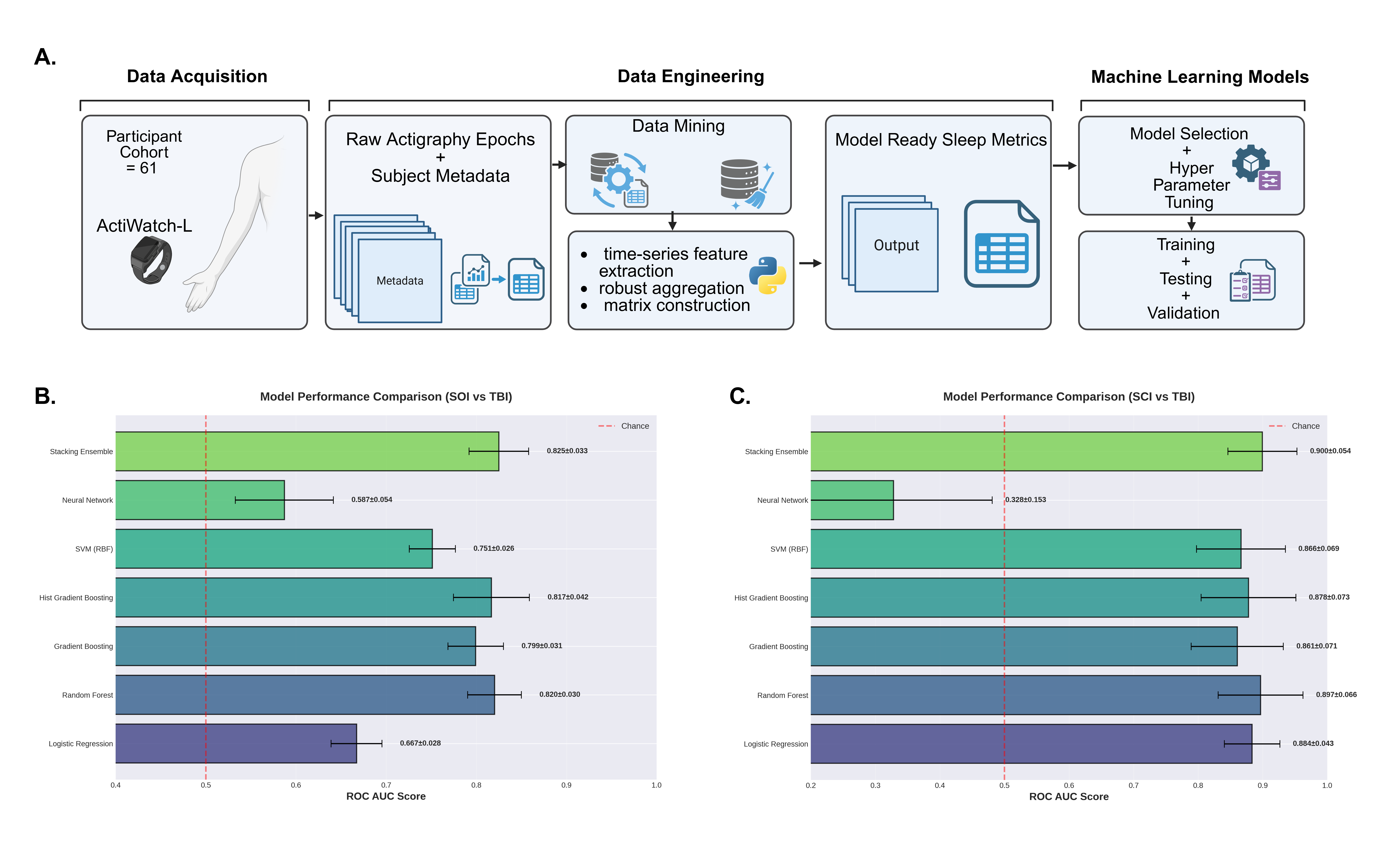 Figure 1. Actigraphy-based sleep feature pipeline and machine learning model performance for TBI classification.
Figure 1. Actigraphy-based sleep feature pipeline and machine learning model performance for TBI classification.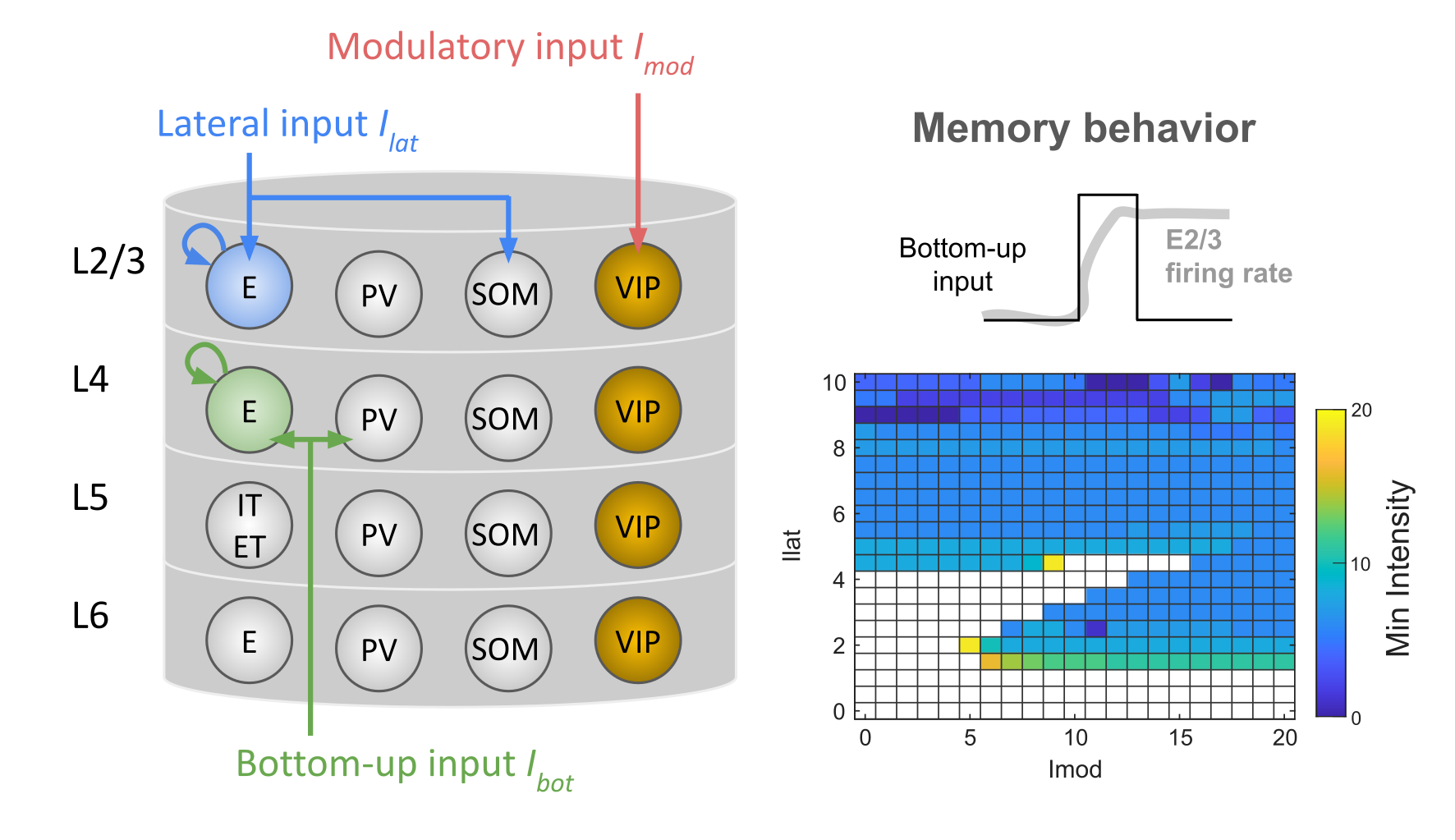 Figure 1. The cortical microcircuit model and its memory behavior. Key model parameters include VIP interneuron cell count (yellow), recurrent connectivity within the E2/3 population (blue), and recurrent connectivity within the E4 population (green). The minimum bottom-up input intensity for a memory behavior varies across different levels of lateral and modulatory inputs.
Figure 1. The cortical microcircuit model and its memory behavior. Key model parameters include VIP interneuron cell count (yellow), recurrent connectivity within the E2/3 population (blue), and recurrent connectivity within the E4 population (green). The minimum bottom-up input intensity for a memory behavior varies across different levels of lateral and modulatory inputs.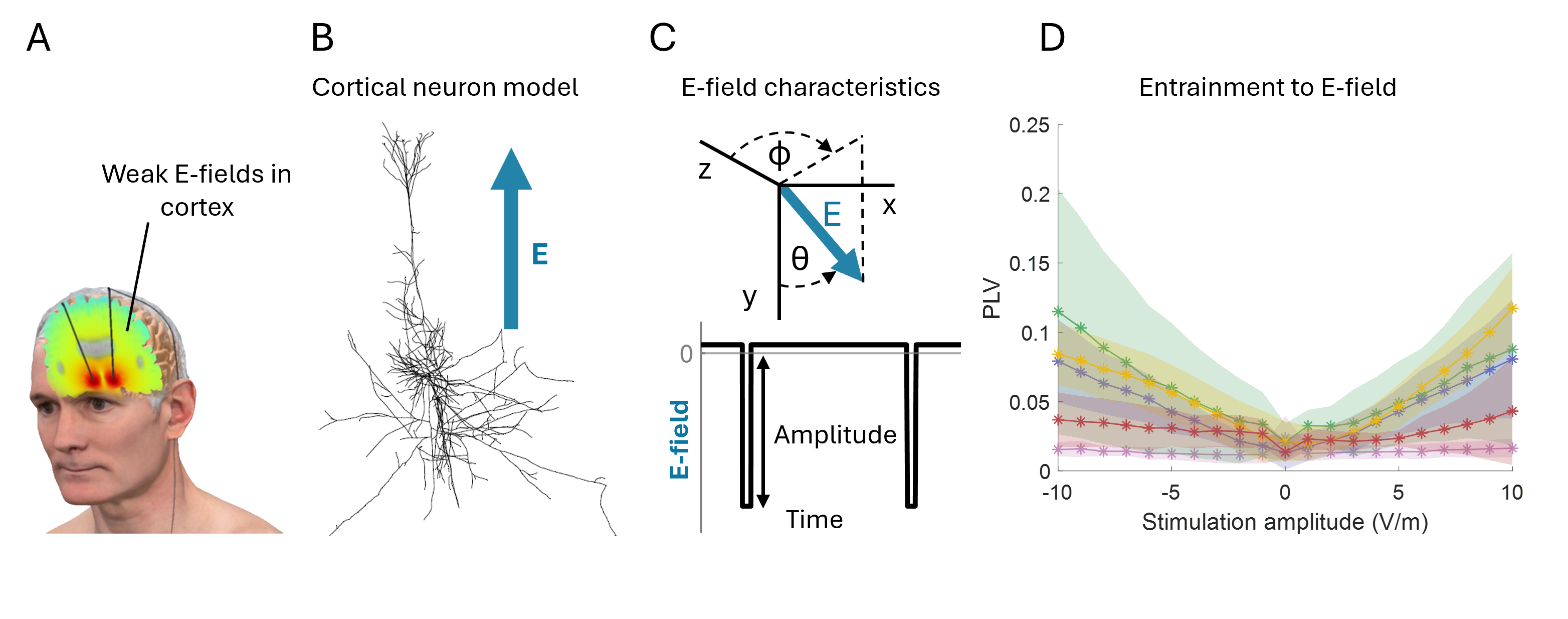 Figure 1. Cortical neuron entrainment by weak electric DBS fields. (A) Illustration of electric fields generated in the brain during DBS. (B) Multi-compartment models of cortical neurons. (C) Electric field orientation relative to the neural morphology. Waveform of the applied field, representing a DBS-like stimulus. (D) Mean phase-locking value increases when amplitude increases.References
Figure 1. Cortical neuron entrainment by weak electric DBS fields. (A) Illustration of electric fields generated in the brain during DBS. (B) Multi-compartment models of cortical neurons. (C) Electric field orientation relative to the neural morphology. Waveform of the applied field, representing a DBS-like stimulus. (D) Mean phase-locking value increases when amplitude increases.References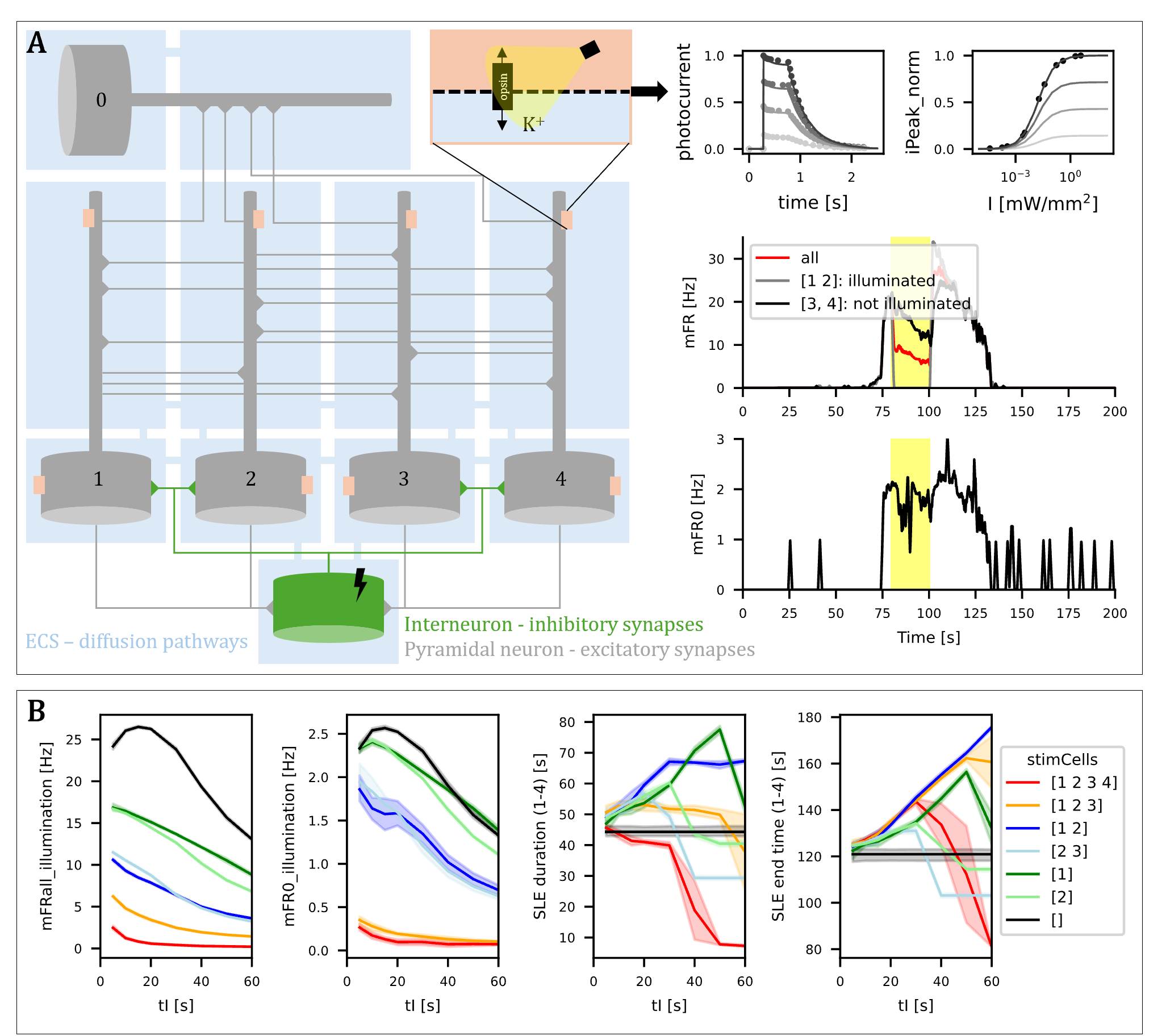 Figure 1. A. Schematic representation of the model and example output. B. Impact of illumination duration (tI) and targeted cell subsets (stimCells) on firing rates during illumination (mFR_all, mFR0) and on SLE duration and end time.
Figure 1. A. Schematic representation of the model and example output. B. Impact of illumination duration (tI) and targeted cell subsets (stimCells) on firing rates during illumination (mFR_all, mFR0) and on SLE duration and end time.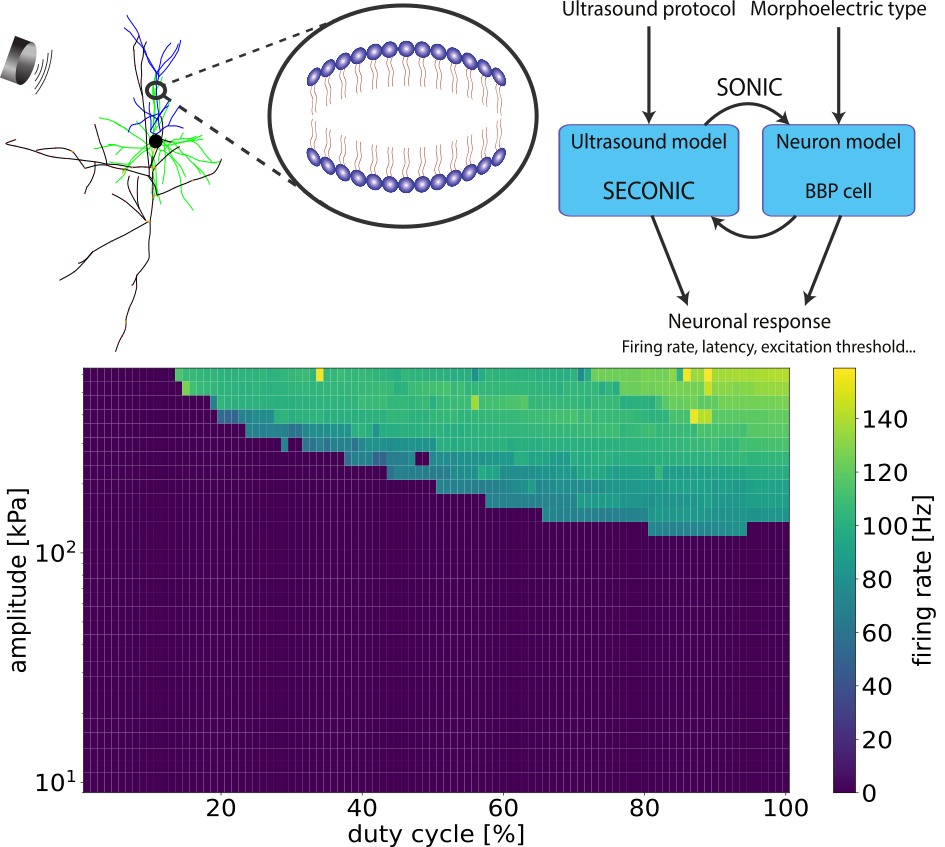 Figure 1. The intramembrane cavitation model in a pyramidal cell (top left), a schematic of the implementation of ultrasound field – neuron coupling (top right) and the firing rate of a L2/3 pyramidal cell as a function of amplitude and duty cycle.
Figure 1. The intramembrane cavitation model in a pyramidal cell (top left), a schematic of the implementation of ultrasound field – neuron coupling (top right) and the firing rate of a L2/3 pyramidal cell as a function of amplitude and duty cycle.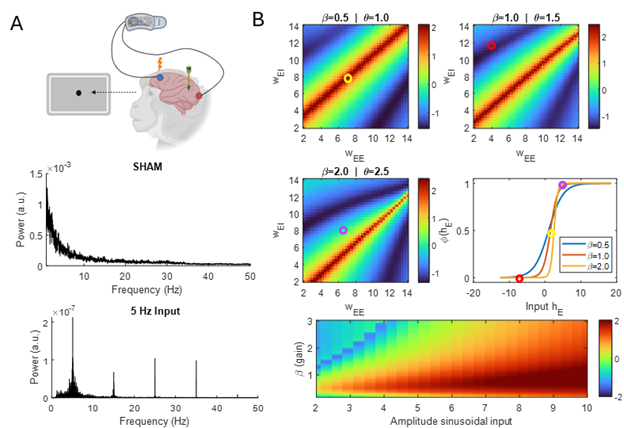 Figure 1. (A) Power spectrum of experimental recordings during SHAM and 5 Hz sinusoidal stimulation. Stimulation induces odd harmonics (15–35 Hz). (B) Log10-OEHR heatmaps from the Wilson–Cowan model showing harmonic dominance across recurrent connectivity (red = odd, dark blue = even). Connectivity sets the net input to the sigmoid transfer function and thus the operating regime (circles).References
Figure 1. (A) Power spectrum of experimental recordings during SHAM and 5 Hz sinusoidal stimulation. Stimulation induces odd harmonics (15–35 Hz). (B) Log10-OEHR heatmaps from the Wilson–Cowan model showing harmonic dominance across recurrent connectivity (red = odd, dark blue = even). Connectivity sets the net input to the sigmoid transfer function and thus the operating regime (circles).References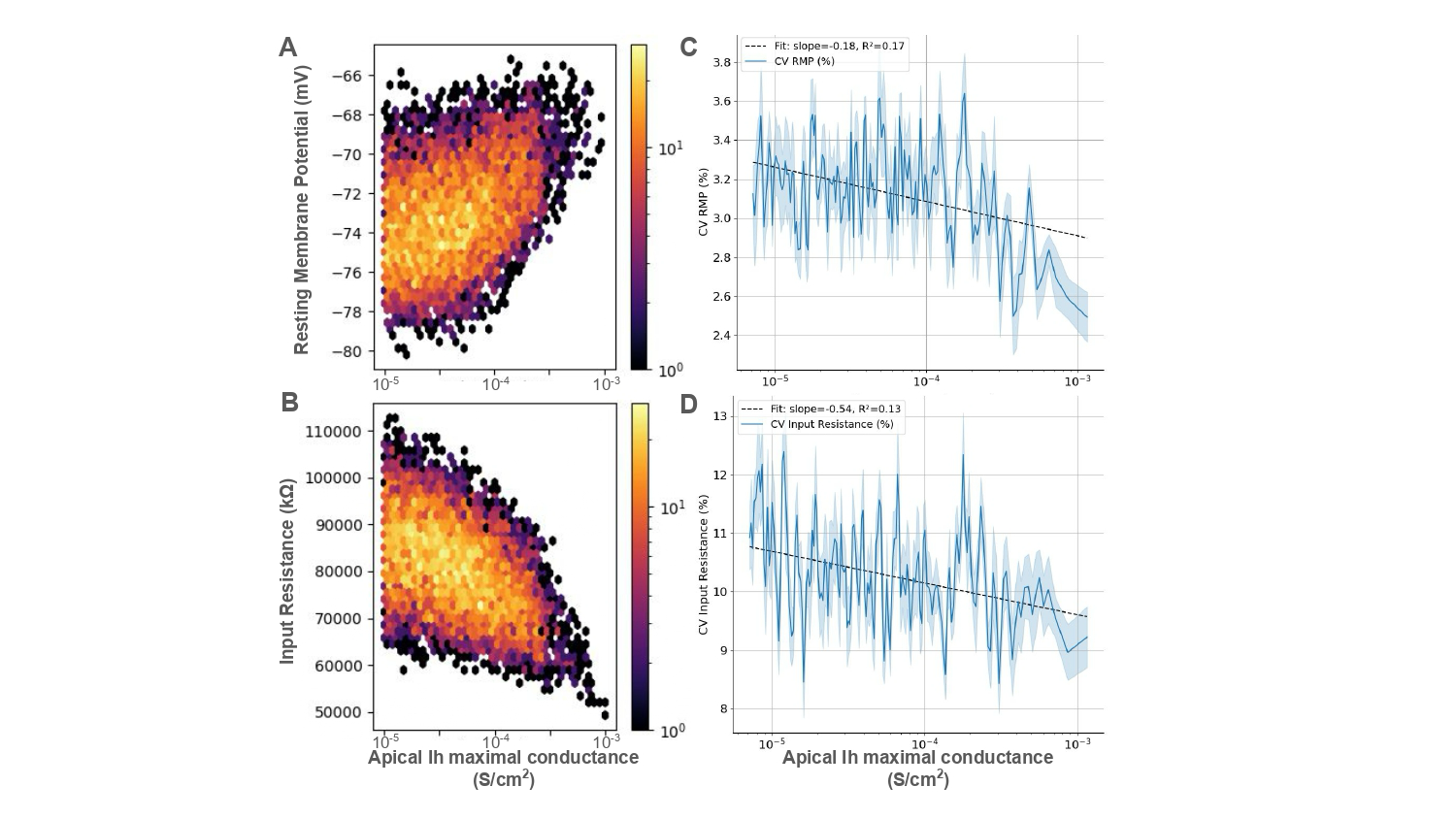 Figure 1. Scatter plot of the (A) resting membrane potential (RMP) and (B) input resistance of a population of ~8000 neurons sorted in increasing order of apical H-current (Ih) maximal conductance, with the coefficient of variation (CV) of RMP (C) and input resistance (D) respectively plotted on the right.
Figure 1. Scatter plot of the (A) resting membrane potential (RMP) and (B) input resistance of a population of ~8000 neurons sorted in increasing order of apical H-current (Ih) maximal conductance, with the coefficient of variation (CV) of RMP (C) and input resistance (D) respectively plotted on the right.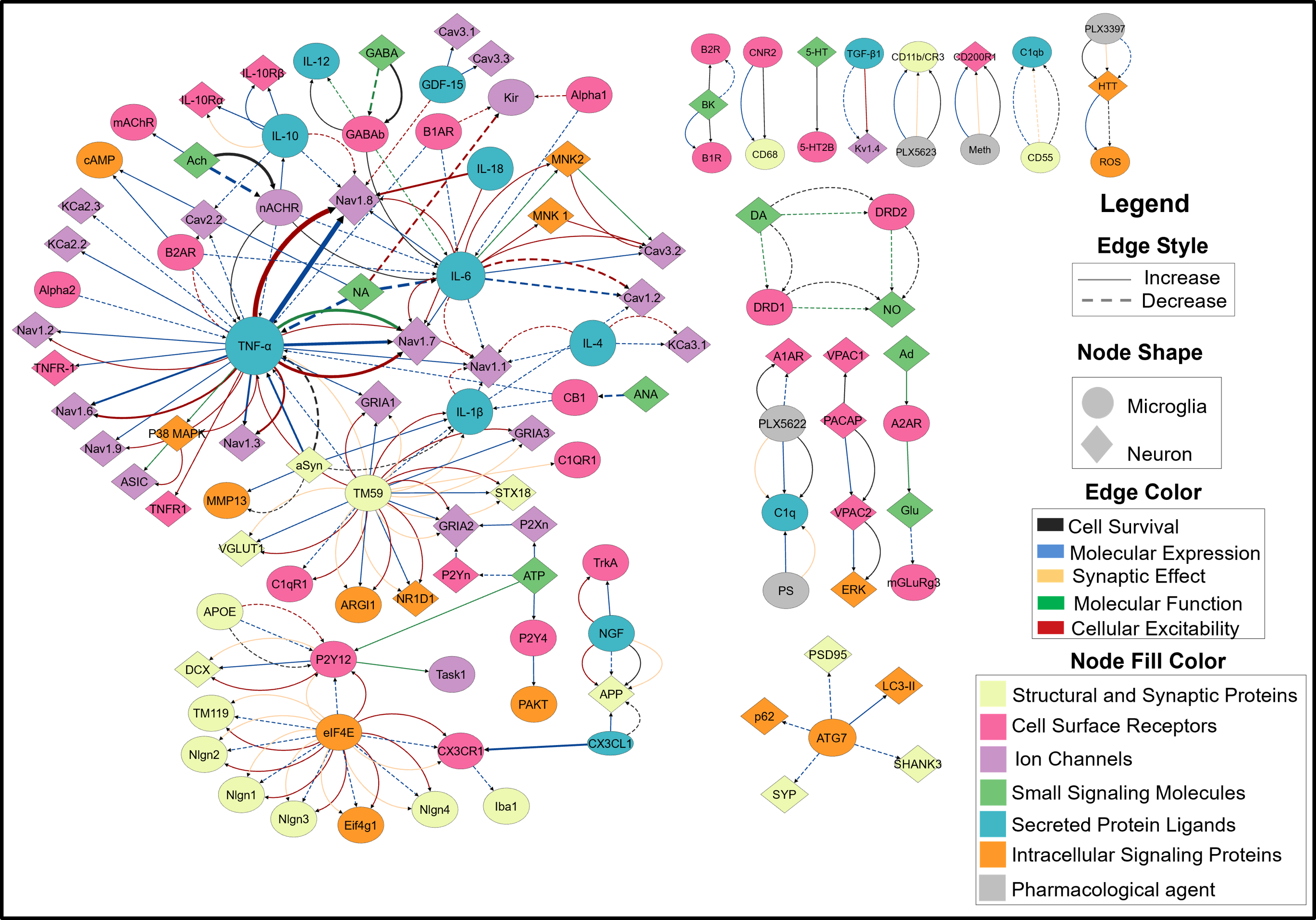 Figure 1. A FINS model of neuroimmune crosstalk. Nodes represent biomolecules, node shape indicates the cell type in which one or more studies reported its localization. The node size corresponds to the number of PRAs in which the node was studied. The node color correspond to the functional category assigned in the nervous system. Edge represents pairwise interaction. Their color shows interaction types, aReferences
Figure 1. A FINS model of neuroimmune crosstalk. Nodes represent biomolecules, node shape indicates the cell type in which one or more studies reported its localization. The node size corresponds to the number of PRAs in which the node was studied. The node color correspond to the functional category assigned in the nervous system. Edge represents pairwise interaction. Their color shows interaction types, aReferences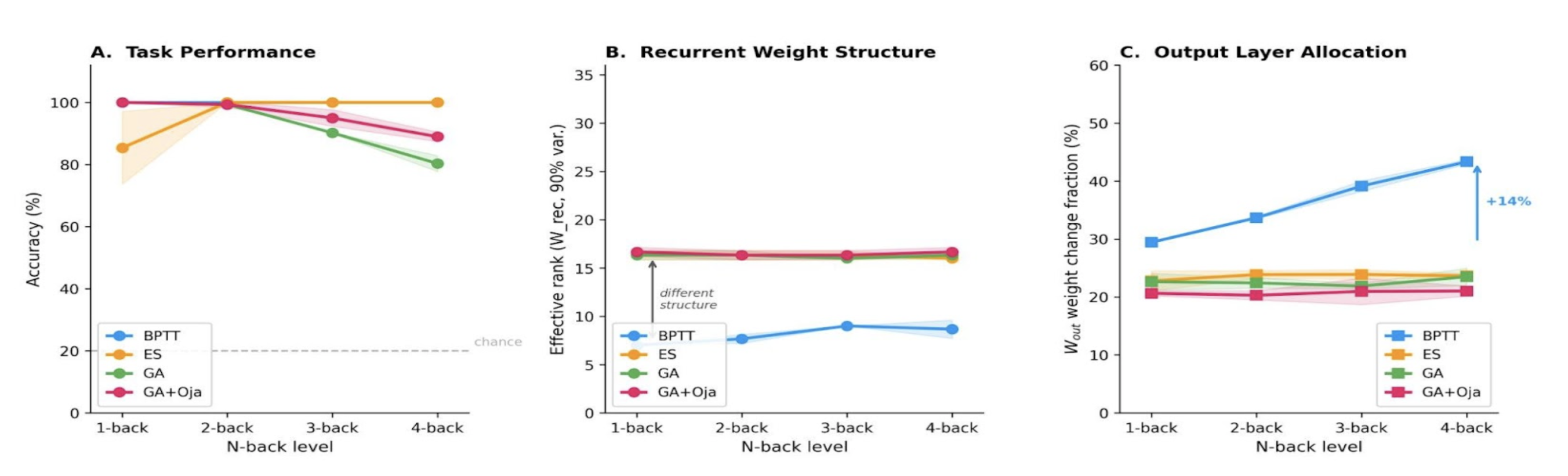 Figure 1. (A) Accuracy vs. n-back level: methods converge at 2-back; ES maintains ~100% at 4-back, whereas GA declines to ~80%. (B) BPTT effective rank increases from ~8 to ~26 with difficulty; evolutionary methods cluster at ~16. (C) BPTT shifts learning to the output layer (+14%); evolutionary methods stay flat. 32 neurons, mean ± std, 3 seeds.
Figure 1. (A) Accuracy vs. n-back level: methods converge at 2-back; ES maintains ~100% at 4-back, whereas GA declines to ~80%. (B) BPTT effective rank increases from ~8 to ~26 with difficulty; evolutionary methods cluster at ~16. (C) BPTT shifts learning to the output layer (+14%); evolutionary methods stay flat. 32 neurons, mean ± std, 3 seeds.



