Cecilia Romaro*
1, Matei Coldea
2, William W. Lytton
3,4, and Robert A. McDougal
1,5,6,7,81 Department of Biostatistics, Yale School of Public Health, New Haven, CT, United States
2 Yale College, Yale University, New Haven, CT, United States
3 Department of Physiology and Pharmacology & Neurology, SUNY Downstate Health Sciences University, Brooklyn, New York
4 Department of Neurology, Kings County Hospital Center, Brooklyn, New York
5 Department of Biomedical Informatics and Data Science, Yale School of Medicine, New Haven, CT, United States
6 Program in Computational Biology and Bioinformatics, Yale University, New Haven, CT, United States
7 Wu Tsai Institute, Yale University, New Haven, CT, United States
8 Interdepartmental Neuroscience Program, Yale University, New Haven, CT, United States
* Email:
[email protected]IntroductionJust as neuron morphology influences spiking behavior and thus network interactions, so too does the 3D placement of spines affect interaction between spines [1] and thus cellular behavior. However fine spine details are not visible under the optical microscopy used for reconstructing neuron morphology and full-cell scanning electron microscopy (SEM) images are generally not feasible due to size constraints. To address these challenges, we developed a tool for the NEURON simulator [2] for importing and editing an SEM reconstruction of a portion of a dendrite, selecting spines, rotating them, and inserting them into a full-cell reconstruction for simulation, using our experimental support for reaction-diffusion multigridding in NEURON.MethodsSEM images may be segmented to identify each spine using standard segmentation software then exported to a TIFF stack. We estimate key electrical properties: approximately equivalent length, diameter, volume, and surface area. Our tool loads the image stack and identifies the voxels forming each spine-dendrite boundary so that we can preserve the connection location after transformations. PySide6 is used to provide a graphical interface allowing spines to be selected and manipulated into position; this can also be done programmatically. An algorithm adds/removes voxels to connect the spine cleanly. Transformed spines can be exported to text files for easy editing, enabling iterative refinement.ResultsWe present our graphical tool, examples of relevant data sets, and simulation results. The graphical tool allows visualization of both the loaded SEM data and the placed spines after transformations. The simulations leverage our previous work, allowing a synaptic source (e.g., of IP3) to be placed at a precise 3D location within a spine. We validate the multigrid simulation by comparing to a single unified 3D simulation and contrast it to simplified geometry approximations, illustrating their similarities and differences. In particular, our tool allows toggling between the two representations.DiscussionSupport for imported spine morphologies brings NEURON a step closer to capturing the intricacies of the human brain. The same tool described here can also directly be used for incorporating SEM data of a dendrite as well. It is not feasible to simulate full cells and networks at this level of detail, nor is that necessarily desirable -- simpler models are often more useful for insights -- but our approach allows us to explore localized behavior in detail in a multiscale context with full cell and network simulations. This tool can give us insight on which details model when and allow us to explore detailed biological questions of synaptic plasticity or the role of morphological changes in disease.References1. Huertas, M. A., Newton, A. J., McDougal, R. A., Sacktor, T. C., & Shouval, H. Z. (2022). Conditions for synaptic specificity during the maintenance phase of synaptic plasticity. Eneuro, 9(3). https://doi.org/10.1523/ENEURO.0064-22.2022
2. Hines, M. L., & Carnevale, N. T. (1997). The NEURON simulation environment. Neural computation, 9(6), 1179-1209. https://doi.org/10.1162/neco.1997.9.6.1179
AcknowledgmentsThis research was funded by the National Institute of Mental Health, National Institutes of Health, grant number R01 MH086638. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
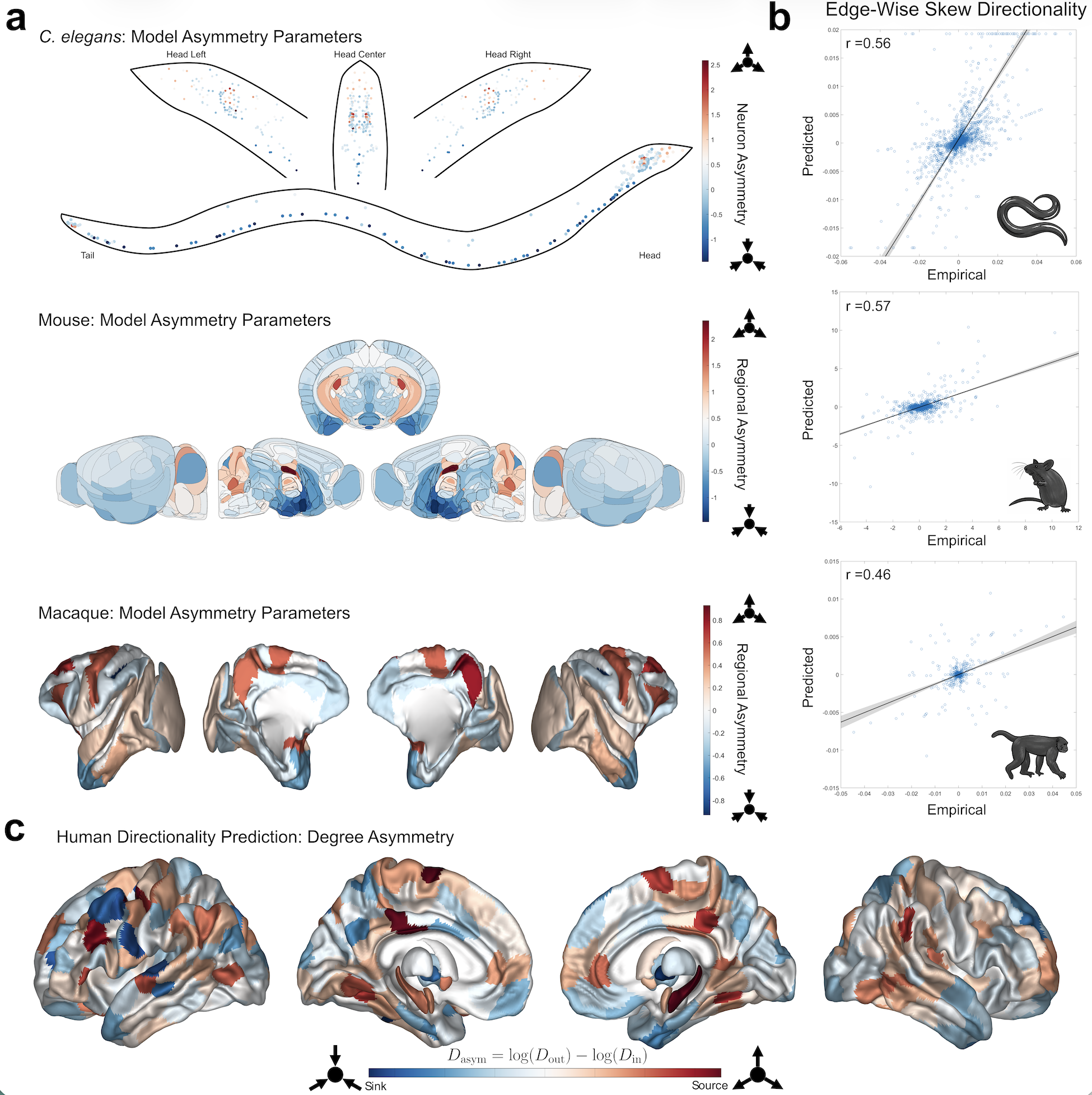 Figure 1. (a) Model-estimated directionality parameters (e^{Ga}) for the three non-human species: C. elegans (top), Mouse (middle), Macaque (bottom). In the C. elegans plot, each dot represents a single neuron. (b) Scatter plots comparing empirical to predicted skew edges with Pearson correlations listed at the top left (all p<10^{-37}). (c) Predicted human overall degree asymmetry for 414 brain regions.
Figure 1. (a) Model-estimated directionality parameters (e^{Ga}) for the three non-human species: C. elegans (top), Mouse (middle), Macaque (bottom). In the C. elegans plot, each dot represents a single neuron. (b) Scatter plots comparing empirical to predicted skew edges with Pearson correlations listed at the top left (all p<10^{-37}). (c) Predicted human overall degree asymmetry for 414 brain regions.
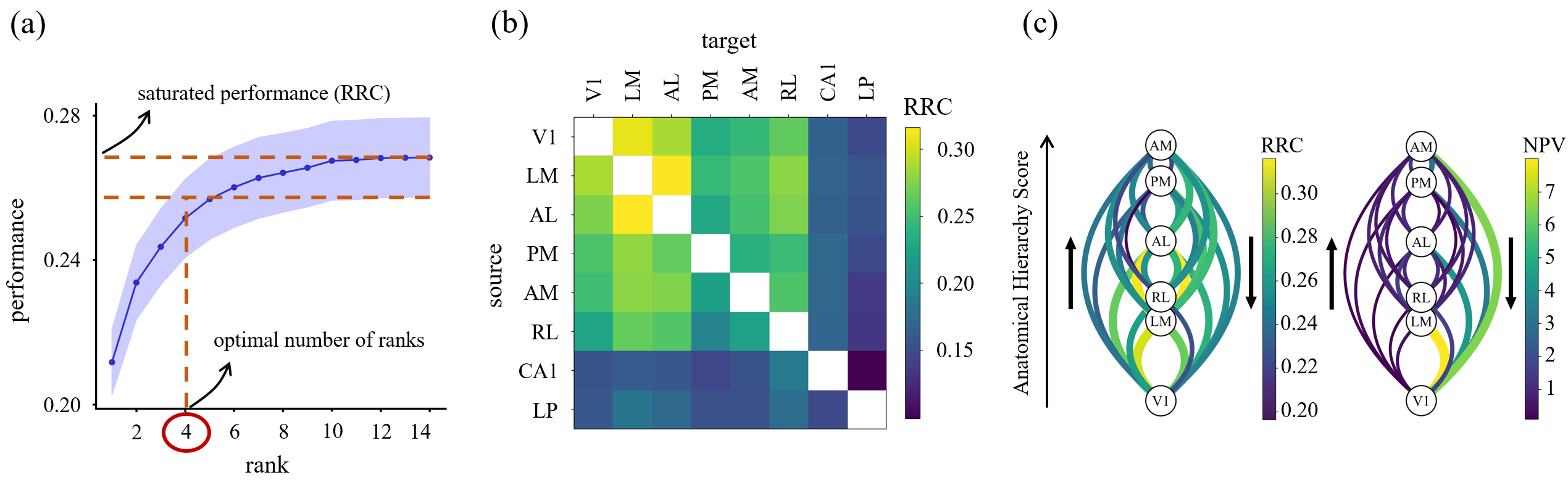 Figure 1. Low-dimensional communication subspaces define RRC. (a) Prediction performance vs. rank; saturation defines optimal number of ranks and RRC. (b) Average RRC across animals; cortical areas are more predictable than subc
Figure 1. Low-dimensional communication subspaces define RRC. (a) Prediction performance vs. rank; saturation defines optimal number of ranks and RRC. (b) Average RRC across animals; cortical areas are more predictable than subc
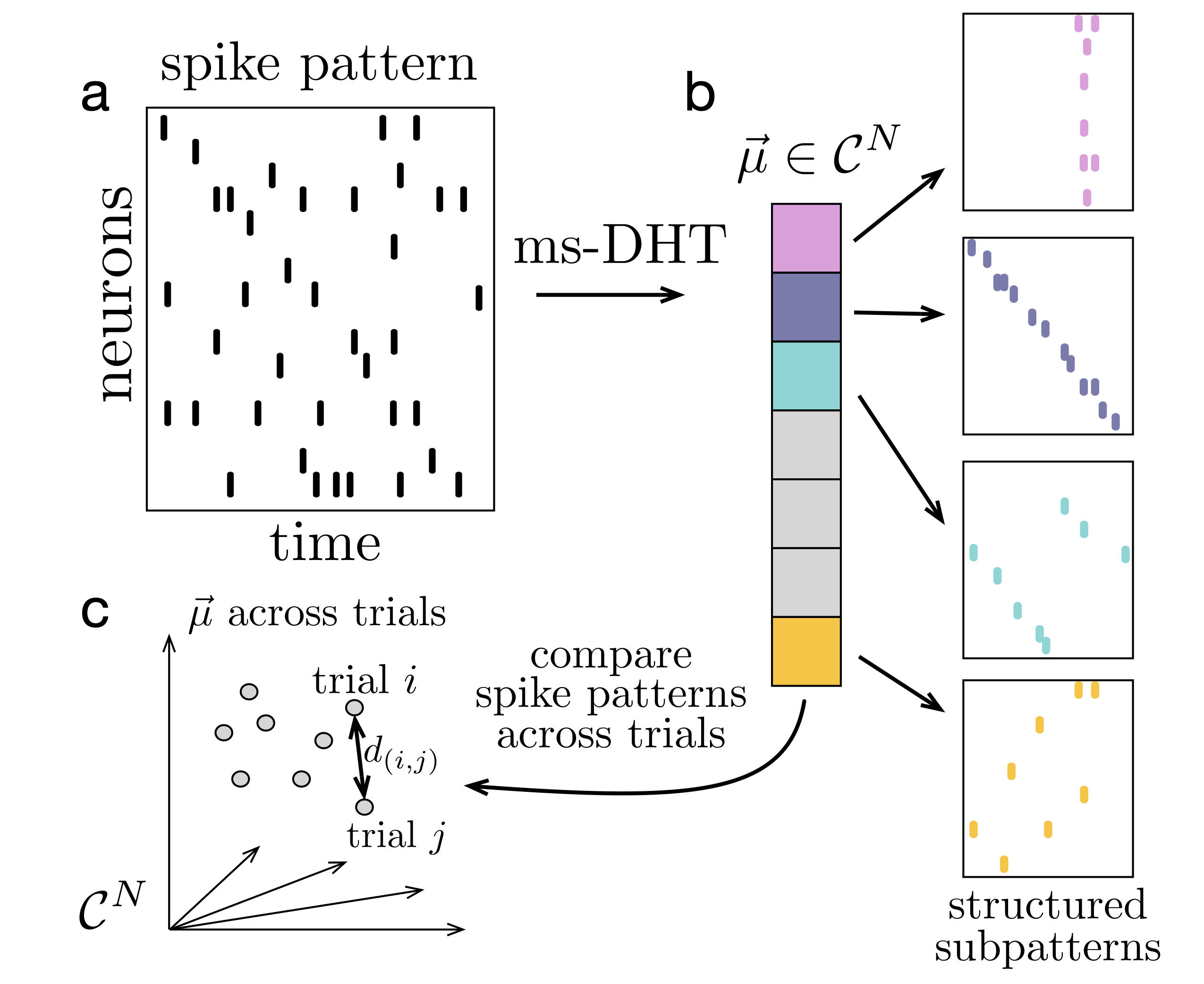 FIgure 1. Decomposing spike patterns. The ms-DHT maps a spike pattern (a) to a unique complex-valued vector (b). Each component encodes the strength (amplitude) and timing (phase) of a basis sub-pattern. (c) Distances between spike patterns reduce to Euclidean distances between ms-DHT outputs, which are invariant to neuron order, ensuring behaviourally relevant clusters do not depend on neuron order.
FIgure 1. Decomposing spike patterns. The ms-DHT maps a spike pattern (a) to a unique complex-valued vector (b). Each component encodes the strength (amplitude) and timing (phase) of a basis sub-pattern. (c) Distances between spike patterns reduce to Euclidean distances between ms-DHT outputs, which are invariant to neuron order, ensuring behaviourally relevant clusters do not depend on neuron order. Figure 1. The CL-1 device is scalable desktop device compatible with standard server racks that allows real-time closed-loop interactions with neural cells via an MEA reader. The CL-1 has onboard hardware that interprets simple code via a Python API to allow rapid code development and experimental iterations coupled with a closed-loop perfusion circuit to automatically adjusts gas levels and temperature to
Figure 1. The CL-1 device is scalable desktop device compatible with standard server racks that allows real-time closed-loop interactions with neural cells via an MEA reader. The CL-1 has onboard hardware that interprets simple code via a Python API to allow rapid code development and experimental iterations coupled with a closed-loop perfusion circuit to automatically adjusts gas levels and temperature to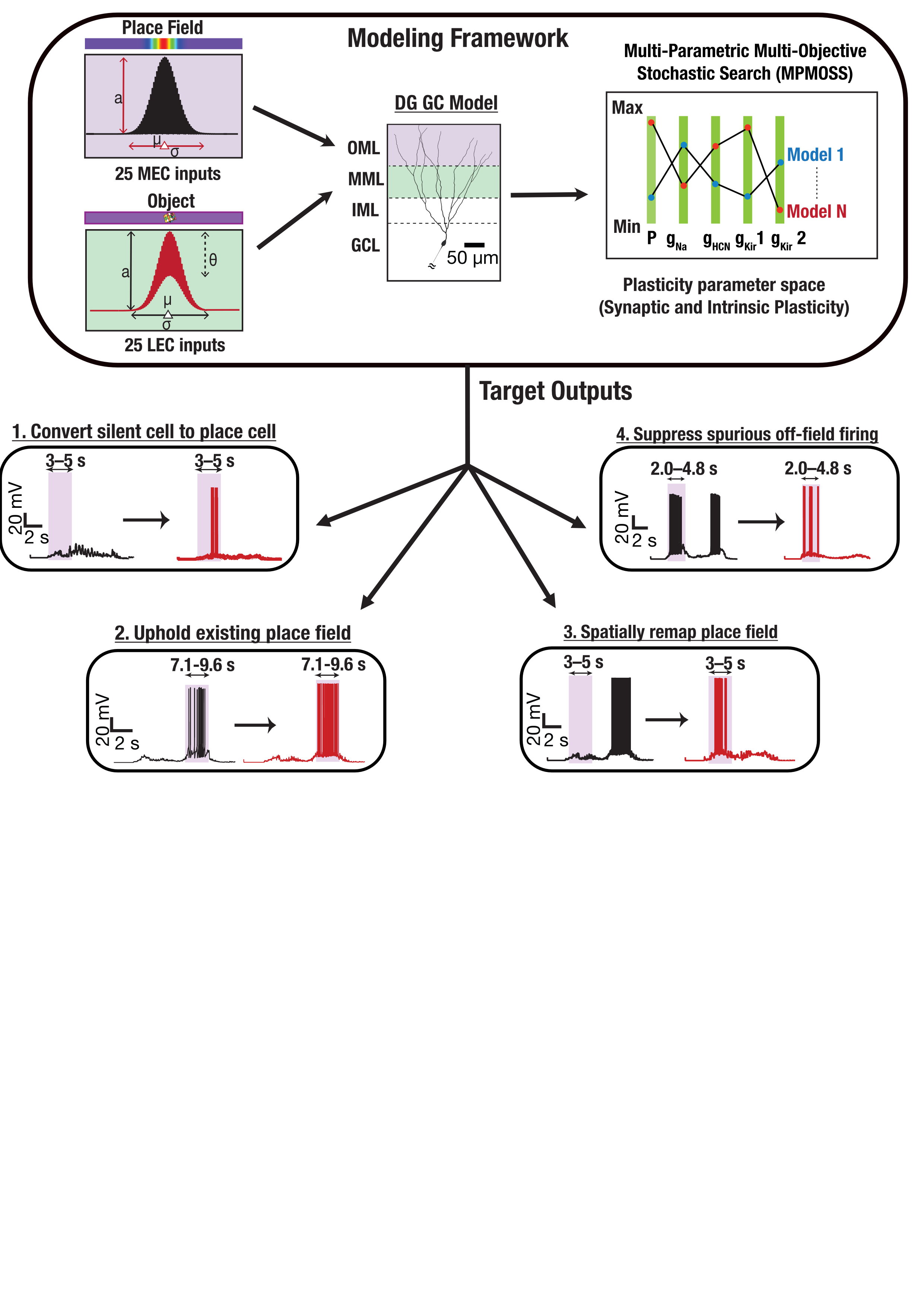 FIgure 1. Medial and lateral entorhinal cortex inputs impinge on a DG granule cell. Disparate combinations of synaptic and intrinsic plasticity (NaP, HCN, Kir channels) achieved one of four targets: convert silent cell to place cell, uphold existing place field, remap, or suppress spurious firing. Our results show that robust and flexible spatial tuning is achievable through plasticity degeneracy.References
FIgure 1. Medial and lateral entorhinal cortex inputs impinge on a DG granule cell. Disparate combinations of synaptic and intrinsic plasticity (NaP, HCN, Kir channels) achieved one of four targets: convert silent cell to place cell, uphold existing place field, remap, or suppress spurious firing. Our results show that robust and flexible spatial tuning is achievable through plasticity degeneracy.References



