IntroductionIt is crucial for an animal’s survival to remember the path taken to reach an important area of a new environment. In the weakly electric fish, neurons in a thalamic-like region respond to encounters with new objects in bursts whose intensity depends on the time elapsed between events, a viable strategy to encode spatial information [1]. It was recently shown that, with such adaptive responses, parametric heterogeneity is necessary to accurately recall a sequence of multiple encounters from the latest response [2]. Here, we generalize the result to include working memory (WM) of past adaptive responses and show that variety in adaptation and threshold parameters maximizes mutual information (MI) between responses and time sequences.
MethodsTiming information is encoded in a resource variable x(n) = 1 - exp(-T(n)/τ)(1-βx(n-1)), where T(n) is the time interval between encounters n-1 and n, τ is the resource recovery rate and β is its history dependence. The response during an encounter n has Poisson statistics with a firing rate proportional to the rectified value max(x-s,0), s being a response threshold (RT) under which a cell does not respond (see Fig 1A). We model WM with a mixture of this distribution and a uniform distribution whose weight increases with the time since the original event (see Fig 1B). This is to represent the unavoidable – and necessary [3] – forgetting of past stimuli. We then use stochastic gradient ascent to optimize MI with respect to the parameters.
ResultsTo quantify the effect of heterogeneity, we look at the number of neurons N necessary to reach 80% of the theoretical MI limit for different numbers of optimized population P and sequence lengths (see Fig 1C). Without WM or RT, the optimized parameters are effectively separated into n different populations. As such, no significant efficiency gain is achieved by increasing P past n. The extra representational capacity of heterogeneous parameters becomes advantageous once WM or RT are available. With or without WM, heterogeneous RT provides a more efficient method to encode time interval sequences, requiring fewer neurons for the same information content. In all cases, heterogeneity offers diminishing returns beyond some value of P.
DiscussionThe cases without RT and WM are consistent with previous results which showed that 1 population is optimal when n=1 and that at least n different populations are necessary for n>1 [2]. WM allows for a middle ground between both cases, as recollections of previous responses are essentially a noisier version of the single interval case with redundancy coming from later responses. Heterogeneous RT allows for division of labor: a population which does not activate indicates time intervals are in the area given by x<s. Such silent coding was shown to increase spatial information in the same fish [4]. This stratagem might also be indicative of the power of rectified nonlinearities commonly observed across neural systems in sequence encoding.
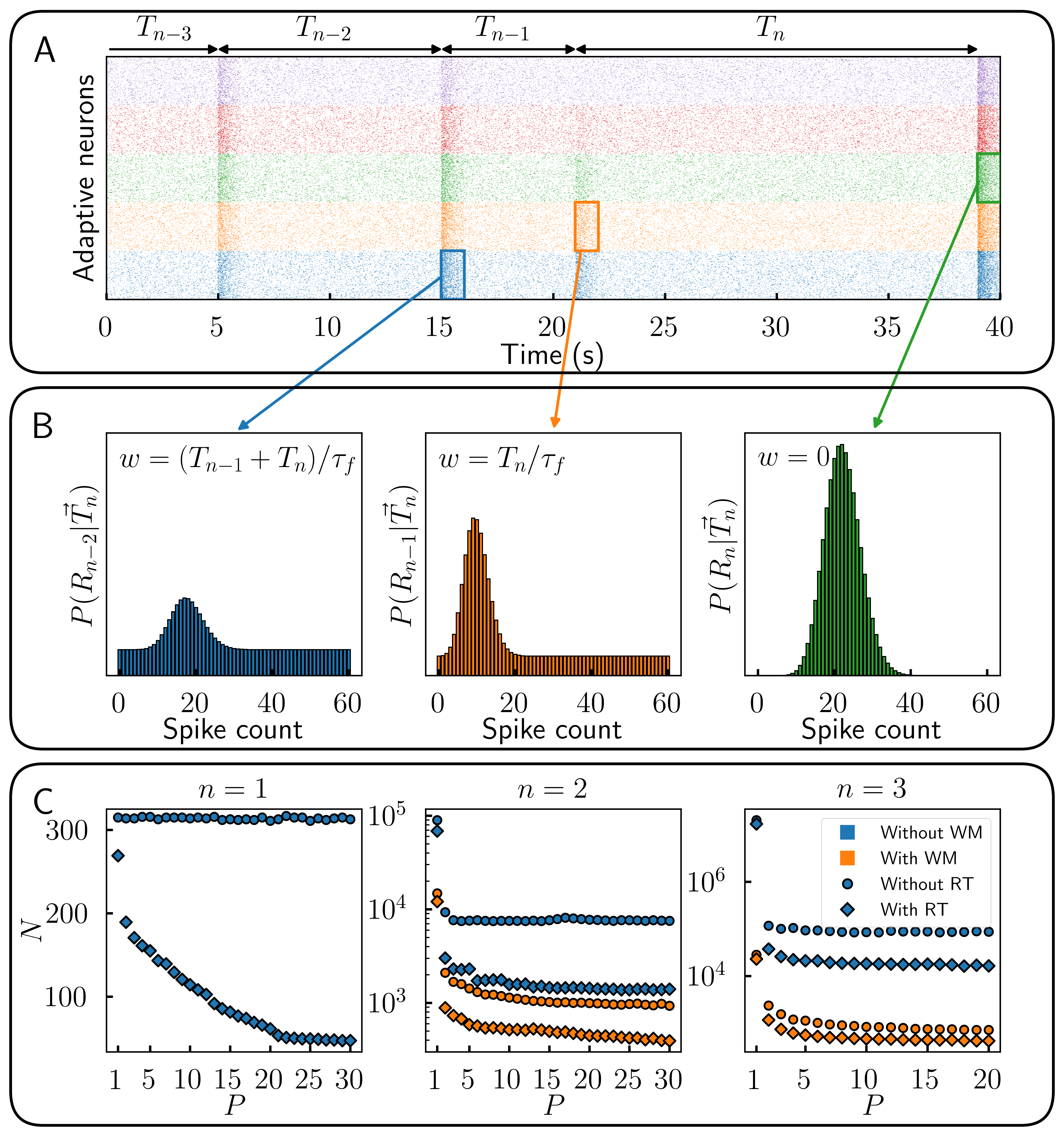 Figure 1. Encoding time sequences is more efficient with parametric heterogeneity. (A) Responses of multiple populations of adaptive cells with response threshold. (B) Recollection of past responses (working memory) during event n. (C) Number of cells N necessary to reach 80% of theoretical maximum information content about a sequence of length n for different number of optimized populations P.
Figure 1. Encoding time sequences is more efficient with parametric heterogeneity. (A) Responses of multiple populations of adaptive cells with response threshold. (B) Recollection of past responses (working memory) during event n. (C) Number of cells N necessary to reach 80% of theoretical maximum information content about a sequence of length n for different number of optimized populations P.
References1. Wallach, A. et al. 2018. A Time-Stamp Mechanism May Provide Temporal Information Necessary for Egocentric to Allocentric Spatial Transformations. eLife 7:e36769. doi:10.7554/eLife.3676
2. Lafond-Mercier, R. et al. 2025. Neural Heterogeneity Enables Adaptive Encoding of Time Sequences. Communications Physics 8(1):504. doi:10.1038/s42005-025-02408-3
3. Georgiou, A. et al. 2021. Retroactive Interference Model of Forgetting. The Journal of Mathematical Neuroscience 11(1):4. doi:10.1186/s13408-021-00102-6
4. Haggard, M., & Chacron, M. J. 2025. Nonresponsive Neurons Improve Population Coding of Object Location. Journal of Neuroscience 45(3). doi:10.1523/JNEUROSCI.1068-24.2024
AcknowledgementThis work was supported by the Natural Sciences and Engineering Research Council of Canada (NSERC) RGPIN/06204-2014 (A.L.) and by the Fonds de recherche du Québec – Nature FRQ B2X/328560 (R.L.M.)
