IntroductionDBS alleviates Parkinsonian symptoms at high frequencies (>90 Hz) yet worsens them at low frequencies (<60 Hz) across STN, GPi, VIM, and SNr; no mechanism explains why frequency alone reverses outcome [1,2]. Pathological beta-band (13–30 Hz) synchronisation in the basal ganglia–thalamocortical loop is the hallmark of Parkinson's disease (PD); its suppression is the leading hypothesis for DBS efficacy [2,3]. High-frequency DBS depresses glutamatergic over GABAergic terminals, shifting E/I balance toward inhibition — an asymmetry attenuated at low frequencies [4,5]. We present a bi-population CANN [6,7] that unifies bistability, beta oscillations, spectral criticality, and spatial responses in a single tractable framework.
MethodsE and I populations sit on a periodic ring with exponential connectivity and ReLU transfer functions [6,7]. DBS is a periodic pulse train; glutamatergic drive scales as F(f) = max(1 + βf, 0), β < 0, the linearised Tsodyks–Markram depression [8]. GABAergic terminals receive a fixed fraction η without attenuation, encoding differential terminal depression [4]. Under uniform stimulation the network reduces to coupled ODEs with Jacobian eigenvalues λ₁,₂ = α ± iω₀ around the active fixed point. Spatial profiles use Fourier decomposition and a Green's function with exact ring boundary conditions. Stochastic fluctuations enter via the linear noise approximation.ResultsAttenuating excitatory drive produces a boundary equilibrium bifurcation at f_th ≈ 90–140 Hz, matching the clinical therapeutic window [1]: below f_th, bistability and hysteresis coexist; above it, excitatory firing is suppressed while inhibitory output grows linearly with frequency, explaining the paradoxical GABAergic increase [4,5]. Beta oscillations emerge when Δ < 0 and α > 0; the Hopf boundary depends only on intrinsic parameters and is invariant to DBS frequency. The spectral criticality index C = ω₀/(2|α|) diverges as α → 0⁻, providing a real-time LFP biomarker for pathological synchrony. Mean-field theory agrees with simulation (N = 100 per population) quantitatively across 1–200 Hz (Fig. 1).
DiscussionDBS therapy and PD pathology act through distinct bifurcations — fully decoupling therapeutic from pathological mechanisms — explaining why DBS is effective without resolving the underlying circuit vulnerability. Testable predictions include: frequency-ramp hysteresis; slope discontinuity in the inhibitory rate–frequency curve at f_th; pre-symptomatic spectral narrowing; nucleus-specific spatial footprints scaling with axonal reach; and stronger pre-operative beta power predicting faster therapeutic onset.
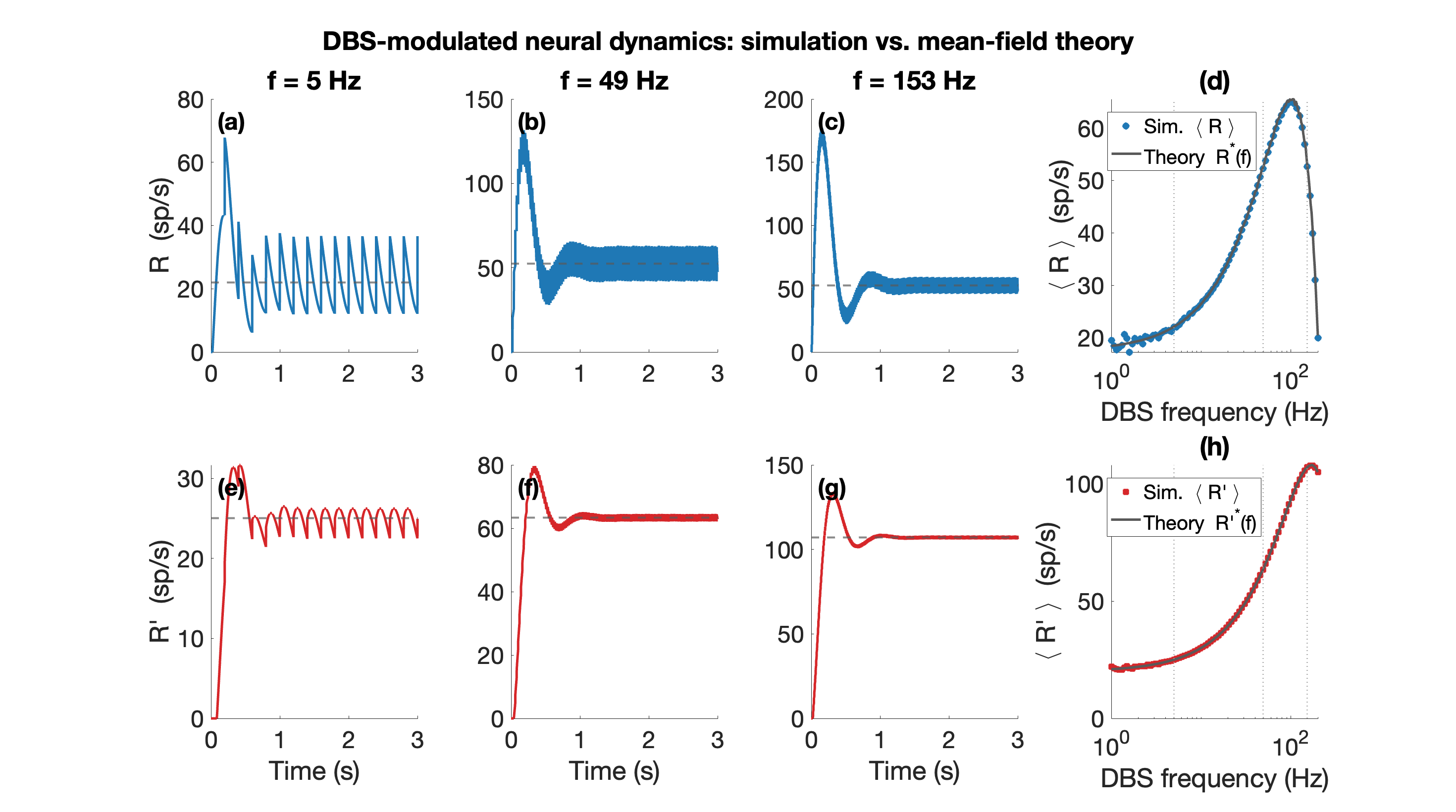 Figure 1. Mean-field theory vs. simulation (N=100 per population). (a)-(c) Excitatory rate R(t) over 0-3 s at f=5, 49, 153 Hz (blue); grey dashed: mean-field prediction; rate fluctuates around mean. (e)-(g) Inhibitory rate R'(t) (red), same convention. (d) Mean R and (h) mean R' at steady state vs. DBS frequency (log scale); circles: simulation; grey line: theory.
Figure 1. Mean-field theory vs. simulation (N=100 per population). (a)-(c) Excitatory rate R(t) over 0-3 s at f=5, 49, 153 Hz (blue); grey dashed: mean-field prediction; rate fluctuates around mean. (e)-(g) Inhibitory rate R'(t) (red), same convention. (d) Mean R and (h) mean R' at steady state vs. DBS frequency (log scale); circles: simulation; grey line: theory.
References[1] McIntyre, C. C., et al. (2004). Clinical Neurophysiology, 115(6), 1239–1248.
[2] Neumann, W.-J., et al. (2023). Brain, 146(11), 4456–4468.
[3] Brittain, J.-S., & Brown, P. (2014). NeuroImage, 85, 637–647.
[4] Li, J., et al. (2025). Nature Neuroscience, 28, 341–355.
[5] Xu, H., et al. (2025). Nature Communications, 16, 245.
[6] Wilson, H. R., & Cowan, J. D. (1972). Biophysical Journal, 12(1), 1–24.
[7] Amari, S. (1977). Biological Cybernetics, 27(2), 77–87.
[8] Tsodyks, M. V., & Markram, H. (1997). PNAS, 94(2), 719–723.
AcknowledgementSupported by the Krembil Brain Institute and the Department of Physiology, University of Toronto. Authors thank colleagues at the Krembil Computational Neuroscience group for discussions.
