IntroductionNeurons can encode information not only by which cells fire, but also by the order in which they fire:
n active neurons can represent
n! sequences. Hippocampal place cells fire in such sequences during navigation and replay these sequences during rest. During replay, synapses can be activated sequentially from tip to soma along dendrites [1]. A dendrite can selectively advance depolarization in response to tip-to-soma inputs, making it
sequence selective [2]. But this selectivity operates within a limited range of AMPA conductance that changes with synaptic spacing, so we ask whether local plasticity can tune these conductances. Our plasticity rule may offer a mechanism for the emergence of plateaus like those observed in BTSP [3].
MethodsVoltage-dependent NMDA and KIR channels make each dendritic segment
bistable: stable at rest and at plateau [4]. A synaptic input briefly opens AMPA channels, depolarizing the segment and potentially transitioning it from rest to plateau. For plateau advancement to be sequence selective, AMPA conductance must be just strong enough that an input transitions a segment from rest to plateau
only with the support of depolarizing current from its plateauing tip-side neighbor. We therefore strengthen AMPA conductance when a segment fails to plateau despite its tip-side neighbor plateauing, and weaken AMPA conductance when it plateaus despite its tip-side neighbor resting. We tested whether this rule operates across different synaptic spacings.
ResultsRepeated tip-to-soma and shuffled input presentations robustly tune each segment’s AMPA conductance into its sequence-selective range. Starting from weak AMPA conductances, analogous to AMPA-silent synapses, inputs initially fail to advance the plateau. Under our plasticity rule, the AMPA conductance of the segment adjacent to the established plateau increases until the segment begins plateauing, after which tuning progresses to the next segment (Fig. 1A). Thus, segments are tuned sequentially from tip to soma, and the number of presentations increases linearly. Because each synapse independently converges on the conductance range it requires, the rule remains effective across heterogeneous synaptic spacings.
DiscussionBecause plateau initiation occurs at a bifurcation, gradual AMPA tuning produces an abrupt transition: a segment initially fails to plateau, then suddenly starts plateauing once AMPA conductance enters the sequence-selective range (Fig. 1B). This sudden plateauing may offer a mechanistic interpretation of BTSP, in which plateaus emerge abruptly after repeated trials: during the initial trials, dendritic plateaus may remain local, but after tuning is complete they may propagate to the soma. Glutamate uncaging experiments could test whether repeated tip-to-soma stimulation selectively strengthens AMPA conductances from tip to soma. Future work should identify a local signal reporting whether the tip-side neighbor plateaued.
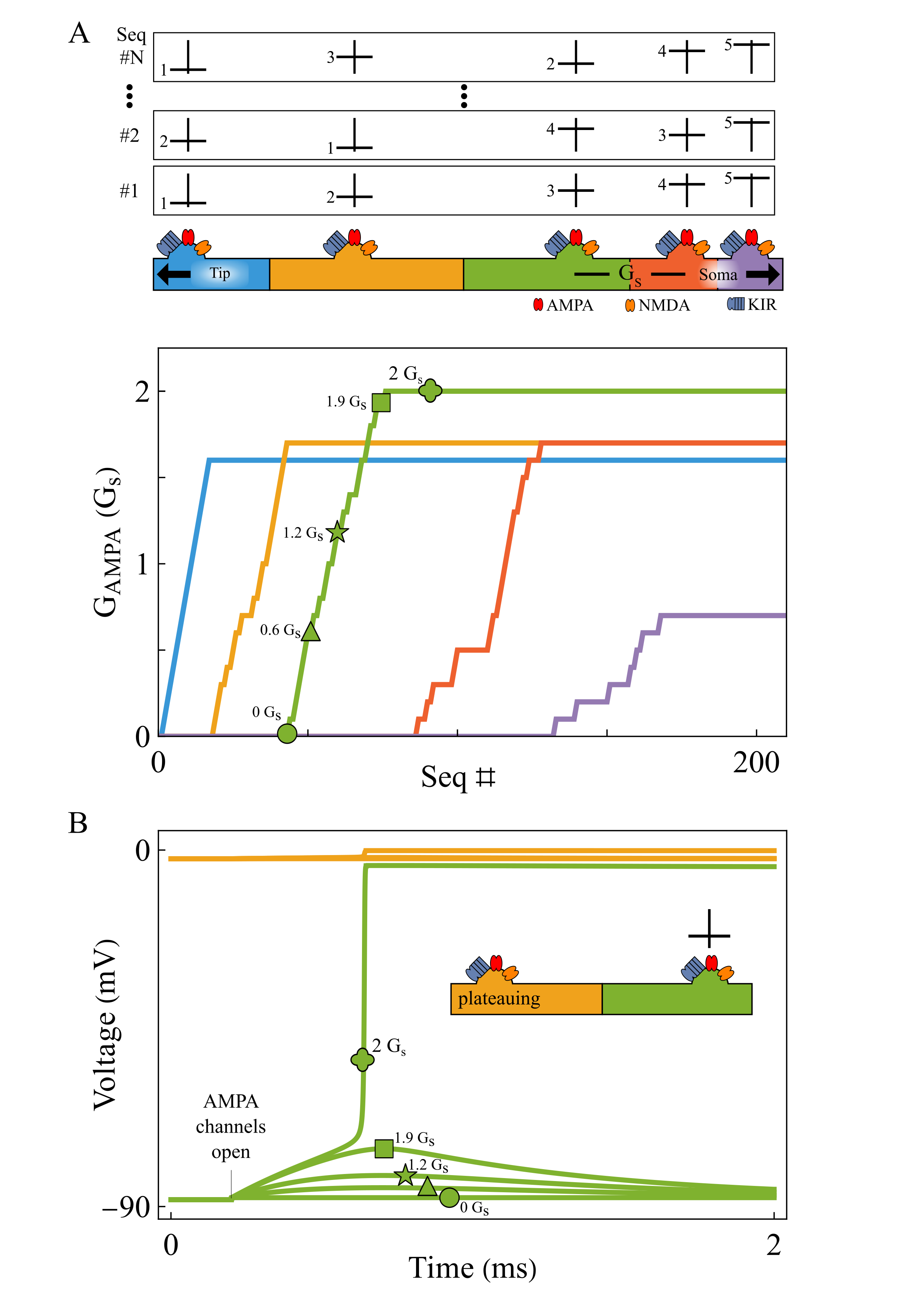 Local AMPA plasticity tunes dendritic sequence selectivity. Repeated tip-to-soma and shuffled inputs progressively strengthen AMPA conductance from tip to soma (A). When a segment (green) receives input while its tip-side neighbor is plateauing (yellow), weak AMPA initially fails to trigger a plateau. As AMPA increases, the segment abruptly transitions to plateau at a bifurcation (B).References
Local AMPA plasticity tunes dendritic sequence selectivity. Repeated tip-to-soma and shuffled inputs progressively strengthen AMPA conductance from tip to soma (A). When a segment (green) receives input while its tip-side neighbor is plateauing (yellow), weak AMPA initially fails to trigger a plateau. As AMPA increases, the segment abruptly transitions to plateau at a bifurcation (B).References- Ishikawa, T., & Ikegaya, Y. (2020). Locally sequential synaptic reactivation during hippocampal ripples. Science Advances, 6(7), Article eaay1492. https://doi.org/10.1126/sciadv.aay1492
- Boahen, K. (2022). Dendrocentric learning for synthetic intelligence. Nature, 612(7938), 43–50. https://doi.org/10.1038/s41586-022-05340-6
- Bittner, K. C., Milstein, A. D., Grienberger, C., Romani, S., & Magee, J. C. (2017). Behavioral time scale synaptic plasticity underlies CA1 place fields. Science, 357(6355), 1033–1036. https://doi.org/10.1126/science.aan3846
- Sanders, H., Berends, M., Major, G., Goldman, M. S., & Lisman, J. E. (2013). NMDA and GABAB (KIR) conductances: The “perfect couple” for bistability. The Journal of Neuroscience, 33(2), 424–429. https://doi.org/10.1523/JNEUROSCI.1854-12.2013
AcknowledgementThis study was supported by the Stanford Bioengineering Department and Masason Foundation.
