Alexandra Busch*,1,2,3, Roberto Budzinski
2,4, Lyle Muller
1,2,31 Department of Mathematics, Western University, London ON, Canada
2 Fields Lab for Network Computation, Fields Lab, Toronto ON, Canada
3 Western Institute for Neuroscience, Western University, London ON, Canada
4 Department of Neuroscience, University of Lethbridge, Lethbridge AB, Canada
*
Email: [email protected]IntroductionRecent technological advances now allow simultaneously recording the activity of thousands of neurons while animals engage in cognitive tasks. These datasets can offer an unprecedented window into how the brain computes in real time, but they also challenge existing analytical frameworks. There has been increasing interest in the possibility that coordinated patterns of spikes, such as sequences, may contribute to neural computation [1-3]. However, in contrast to the many methods available for analyzing firing rates, mathematical tools capable of systematically probing spike-time structure at the scale of these next-generation datasets remain limited.MethodsWe introduce a decomposition operator for population spike patterns, termed the multi-sample Discrete Helix Transform (ms-DHT). We derive a generalized inner product that allows the ms-DHT to operate directly on patterns of discrete spikes across thousands of neurons without smoothing. The ms-DHT decomposes these spike patterns into a fixed, interpretable basis, mapping each input pattern to a unique vector that captures the occurrence and timing of every spike (Fig.1). In this representation, distances between spike patterns reduces to the Euclidean distance between their ms-DHT outputs. This distance is invariant to neuron ordering and allows detecting repeating structure ranging from simple spike sequences to complex population motifs.
ResultsWe demonstrate several applications of the ms-DHT to large-scale datasets. Notably, in dual Utah array recordings from the prefrontal cortex of a macaque monkey performing a virtual reality working memory task, the ms-DHT reveals structured spike motifs that predict specific behavioural errors on single trials - before they occur. Further, applications to spiking network simulations with 10,000 neurons demonstrate that the transform operates effectively at the scale of next-generation neural recordings.DiscussionThe ms-DHT provides a flexible framework for analyzing large-scale spike patterns. By decomposing spiking activity onto a fixed, interpretable basis using a generalized inner product, the ms-DHT produces unique descriptions of population activity even when neurons emit variable numbers of spikes—a setting that has posed a central challenge for analytical approaches. The resulting representation supports multiple analyses, including clustering and decoding of full spike patterns, detecting repeating substructure through specific helix contributions, and sliding-window analyses that trace the temporal evolution of spike patterns across long recordings.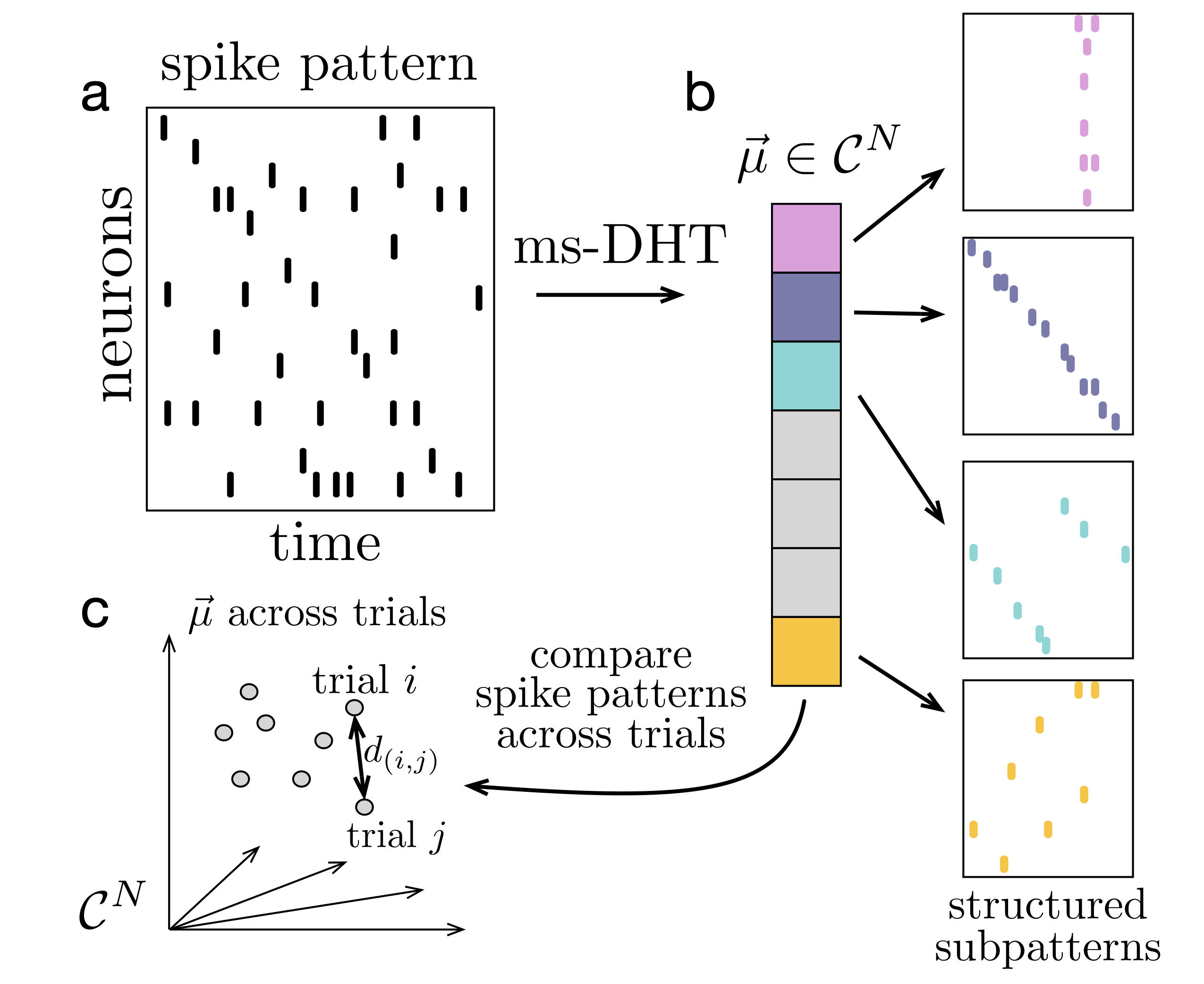 FIgure 1. Decomposing spike patterns. The ms-DHT maps a spike pattern (a) to a unique complex-valued vector (b). Each component encodes the strength (amplitude) and timing (phase) of a basis sub-pattern. (c) Distances between spike patterns reduce to Euclidean distances between ms-DHT outputs, which are invariant to neuron order, ensuring behaviourally relevant clusters do not depend on neuron order.
FIgure 1. Decomposing spike patterns. The ms-DHT maps a spike pattern (a) to a unique complex-valued vector (b). Each component encodes the strength (amplitude) and timing (phase) of a basis sub-pattern. (c) Distances between spike patterns reduce to Euclidean distances between ms-DHT outputs, which are invariant to neuron order, ensuring behaviourally relevant clusters do not depend on neuron order.
References[1] Xie, W., Wittig, J. H., Chapeton, J. I., El-Kalliny, M., Jackson, S. N., Inati, S. K., & Zaghloul, K. A. (2024). Neuronal sequences in population bursts encode information in human cortex. Nature, 635(8040), 935–942. https://doi.org/10.1038/s41586-024-08075-8
[2] Chettih, S. N., Mackevicius, E. L., Hale, S., & Aronov, D. (2024). Barcoding of episodic memories in the hippocampus of a food-caching bird. Cell, 187(8), 1922–1935.e20. https://doi.org/10.1016/j.cell.2024.02.032
[3] Busch, A., Roussy, M., Martinez-Trujillo, J. C., et al. (2024). Neuronal activation sequences in lateral prefrontal cortex encode visuospatial working memory during virtual navigation. Nature Communications, 15, 4471. https://doi.org/10.1038/s41467-024-48664-9AcknowledgmentsThis work was supported by NSERC, CFREF, NIH, Neuronex NSF, and Canada Research Chairs Program. A.B. gratefully acknowledges a BrainsCAN studentship and NSERC CGS-D.
